|
|
Органы, ткани и клетки с эндокринной функциейСтр 1 из 11Следующая ⇒ Кафедра патофизиологии
ОСНОВНЫЕ АСПЕКТЫ ПАТОФИЗИОЛОГИИ ЭНДОКРИННОЙ СИСТЕМЫ Учебное пособие для самостоятельной работы студентов медицинского университета
Рекомендовано Учебно-методическим объединением по медицинскому и фармацевтическому образованию вузов России в качестве учебного пособия для студентов медицинских институтов и университетов. Северьянова Л.А. Основные аспекты патофизиологии эндокринной системы. Учебное пособие для студентов медицинского университета. – Курск: КГМУ, 2006. – 100 с.
Рецензенты: член-корр. РАМН, проф. Г.В. Порядин – зав. кафедрой патофизиологии Российского государственного медицинского университета член-корр. РАМН, проф. А.В. Завьялов – зав. кафедрой нормальной физиологии Курского государственного медицинского университета. Учебное пособие содержит новейшие данные по патофизиологии эндокринной системы, в том числе еще не вошедшие в отечественную учебную литературу, а также собственные разработки автора. Рекомендуется для студентов медицинских и фармацевтических факультетов и для начинающих врачей. Учебное пособие составлено в соответствии с утвержденной Минздравом РФ программой по патофизиологии и действующим учебным планом для высших медицинских учебных заведений.
С О Д Е Р Ж А Н И Е
ГЛАВА 1 ОБЩИЕ ВОПРОСЫ ФИЗИОЛОГИИ ЭНДОКРИННОЙ СИСТЕМЫ
Эндокринная система – специализированная система гуморальной регуляции функций. Гуморальная регуляция – взаимодействие между клетками, тканями, органами и системами органов, осуществляемое биологически активными веществами (БАВ) через жидкие среды организма. Основой эффектов БАВ является избирательная чувствительность к ним клеток-мишеней, которая обусловлена наличием соответствующих рецепторов. В настоящее время сохраняется деление БАВ на следующие группы: гормоны, гистогормоны и парагормоны. Основные типы действия биологически активных веществ на клетки-мишени представлены на рис. 1.
А. Гормоны (истинные гормоны). Важнейшим свойством истинных гормонов считают их образование специализированными популяциями клеток, для которых основной функцией является выделение секрета во внутреннюю среду организма, т.е. функция внутренней секреции – эндокринная функция. При этом выделяют следующие формы организации клеток. 1. Эндокринные железы как целые железистые органы, не имеющие выводных протоков (в отличие от экзокринных желез) и выделяющие гормоны в кровь. 2. Популяции эндокринных клеток, расположенные в различных органах и тканях, для которых внутренняя секреция не является единственной функцией. К этой группе относятся эндокринные клетки головного мозга, в первую очередь гипоталамические нейроны, выделяющие рилизинг-факторы и нейрогормоны, и гастроинтестинальная система, имеющие общее происхождение – из нейроэктодермы. Многие гормоны этой группы относят к регуляторным пептидам.
Свойства истинных гормонов 1. Специфичность действия, проявляющаяся в том, что каждый гормон вызывает строго определенный, характерный только для него эффект. 2. Высокая биологическая активность, проявлением которой служит эффективность чрезвычайно низких концентраций гормона в плазме крови. 3. Выделение в циркулирующие жидкости организма и дистантный характер действия.
Б. Гистогормоны (тканевые гормоны). Имеют следующие характеристики: 1. Образуются в различных органах и тканях. 2. Осуществляют кратковременное действие, т.к. являются короткоживущими соединениями. 3. Действуют локально через межклеточные контакты (паракринное действие) и гуморально в пределах ткани, которая их продуцирует, или на близлежащие ткани. Таким образом, гистогормоны обеспечивают быструю саморегуляцию тканевых процессов. Среди гистогормонов выделяют, как правило, две подгруппы:
Ø Тканенеспецифические, обладающие более широким спектром физиологического действия: вазоактивные кинины, биогенные амины (серотонин, гистамин, аденозин), гепарин и др. Ø Тканеспецифические, отличающиеся более высокой специализацией местного действия: тканеспецифические факторы роста (эпидермиса, фибро-
Рис. 1. Типы действий биологически активных веществ на клетки-мишени. В. Парагормоны – биологически активные метаболиты, не имеющие специфического происхождения, т.е. образующиеся в различных тканях и органах. Они могут приобретать роль специфических внешних регуляторов в отношении различных функций: например, углекислота активирует нейроны бульбарного дыхательного центра, ионы кальция участвуют в регуляции секреторного процесса в железистых клетках и выделения медиаторов нервными окончаниями.
В настоящее время наиболее общепринятым является следующий перечень эндокринных желез и органов с инкреторной функцией (табл. 1).
Таблица 1 I. Биосинтез гормонов Известны два основных типа генетического контроля синтеза гормонов: 1. Прямой контроль – синтез в полисомах предшественников белково-пептидных гормонов, характеризующийся схемой: ген → мРНК → прогормон → гормон. 2. Опосредованный контроль по схеме: гены → мРНК → фермент → гормон. Этой схеме соответствует нерибосомальный синтез стероидных гормонов и производных аминокислот. В соответствии с химическим строением гормоны разделяют в настоящее время на 4 группы: · белки и пептиды; · производные аминокислот; · стероиды; · простагландины.
II. Секреция Секреция многих гормонов осуществляется импульсно, дискретными порциями. При этом секреторный процесс может включать различные механизмы. 1. Освобождение гормона из секреторных гранул, происходящее с использованием энергии АТФ и при участии ионов кальция (белково-пептидные гормоны, катехоламины). 2. Освобождение из белковосвязанной формы (тиреоидные гормоны). 3. Экзоцитоз (эмиоциоз) по схеме: миграция гранул, содержащих гормон, к мембране клетки → растворение мембраны в месте контакта → прорыв содержимого гранулы в межклеточное пространство → диффузия в капилляры. 4. Диффузия через плазматические мембраны (стероидные гормоны). V. Инактивация гормонов В этом конечном этапе любой эндокринной функции принимают участие обменные процессы, протекающие при использовании гормона в реагирующих тканях (активация, реактивация, взаимопревращения), т.е. периферические превращения и катаболизм, а также специальные процессы инактивации, происходящие для многих гормонов в печени (катаболизм стероидных гормонов, дейодирование тиреоидных гормонов и образование эфиров с глюкуроновой и серной кислотой и т.д.). Некоторые гормоны (растворимые в воде), например стероидные, выделяются почками. Для нормального состояния эндокринной функции, естественно, характерно равновесие процессов продукции и инактивации гормона. Эндокринные ритмы
Как и другие организованные функции животных, активность эндокринных желез является объектом периодических и циклических изменений. Некоторые периодические колебания возникают под влиянием внутреннего механизма независимо от внешнего окружения, некоторые – могут быть координированы внешними сигналами, такими, как смена свет-темнота, день-ночь и др. Многие эндогенные ритмы имеют период приблизительно 24 часа (циркадные). Ритмы, которые встречаются чаще, чем 1 раз в сутки, относятся к ультрарадианным. Некоторые периодические явления имеют большую продолжительность (месяц, год и пр.) Циркадные ритмы характерны для большинства эндокринных функций. Для человеческого гипофиза секреция ГР и пролактина является максимальной после засыпания, для кортизола – между 2 и 4 часами утра. Секреция ТТГ самая низкая утром между 9 и 12 часами и максимальная – между 20 и 24 часами. Секреция гонадотропина у развивающихся подростков максимальная ночью. На циркадный ритм накладываются ультрарадианные вспышки гормональной секреции – пульсаторные выделения. Так, секреция гонадотропинов в юности характеризуется быстрыми высокоамплитудными пульсациями, особенно ночью, в то время как у сексуально зрелых людей эпизоды секреции более низкой амплитуды и встречаются в течение 24 часов. Пролактин, гормон роста и АКТГ также секретируются короткими и довольно регулярными импульсами. Что касается механизмов формирования эндокринных ритмов, то считается общепризнанной их регуляция мозгом. Известной нейрональной структурой, ответственной за циркадные ритмы у высших животных, является супрахиазматическое ядро переднего гипоталамуса. Оно имеет ретино-гипоталамический вход (обеспечивающий изменения "свет-темнота") и содержит пептидергические нейроны, продуцирующие соматостатин, ВИП, нейротензин, нейро- Существование эндокринных ритмов имеет важное биологическое значение, так как обеспечивает наиболее адекватную потребностям организма активность органов-мишеней, а также адаптивное поведение. Если бы гормон присутствовал в крови в постоянной достаточно высокой концентрации, клетки-мишени могли бы становиться рефрактерными к нему из-за уменьшения числа свободных рецепторов. Пульсаторное же выделение гормона обеспечивает возможность увеличения частоты импульсов и усиление секреторного ответа клетки-мишени.
ГЛАВА 2 Применение в медицине На базе регуляторных пептидов созданы первые лекарственные препараты. Так, олигопептиды (короткие пептиды) N-терминального фрагмента АКТГ и МСГ используются для коррекции внимания и запоминания, вазопрессин – для улучшения памяти при травматической и других амнезиях. Широкое применение в лечебной практике имеет отечественный препарат даларгин (аналог лей-энкефалина). Начат коммерческий выпуск сурфагона (аналог люлиберина), предназначенного для лечения нарушений репродуктивной системы.
ГЛАВА 3 ГЛАВА 4 ПАТОФИЗИОЛОГИЯ ГИПОФИЗА Эффекты АКТГ 1. Адренокортикотропное действие, т.е. стимуляция секреции коры надпочечников. Основной эффект – активация пучковой зоны и увеличение продукции кортизола, в меньшей степени – сетчатой зоны и секреции андрогенов и в малой степени – альдостерона.
2. Метаболическое действие. В основном зависит от продукции глюкокортикоидов, но включает и вненадпочечниковые эффекты. Изменение белкового обмена при гиперпродукции АКТГ: АКТГ → усиление синтеза кортизола в надпочечниках → усиление катаболизма белков → в условиях повышенного распада белков – усиление анаболизма. Влияние избытка АКТГ на жировой обмен: а) АКТГ → вненадпочечниковый механизм: усиление липолитической активности жировой ткани → усиление распада жиров → выход в кровь свободных жирных кислот и их окисление до кетоновых тел; б) АКТГ → надпочечниковый механизм: торможение мобилизации жира + торможение липолитического действия СТГ → активация глюконеогенеза → стимуляция образования жира.
3. Нейротропное действие обусловлено N-концевым фрагментом молекулы гормона – АКТГ4-10 (4-10 – обозначение последовательности аминокислотных остатков). Основные эффекты: улучшение внимания и кратковременной памяти, ускорение обучения человека и животных, улучшение выбора биологически целесообразного поведения при стрессе.
4. Влияние на образование меланина и пигментацию кожи. Действие аналогично эффекту МСГ и объясняется тем, что АКТГ содержит в своей молекуле всю аминокислотную последовательность меланоцитстимулирующего гормона. Патология
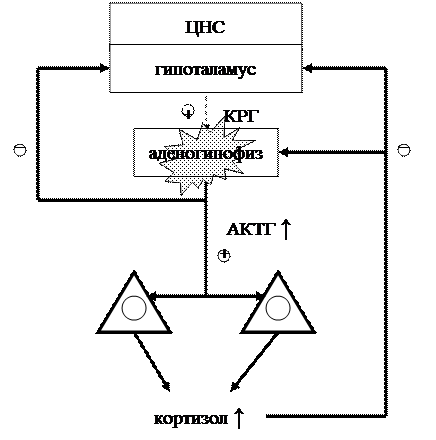
Гиперсекреция АКТГ наблюдается при базофильной аденоме передней доли гипофиза и нарушениях в гипоталамусе (недостаток дофамина) и приводит к развитию болезни Иценко-Кушинга. Основной механизм патогенеза – избыточная стимуляция под действием АКТГ коры надпочечников и развитие вторичного гиперкортизолизма. Гиперпродукция кортизола проявляется характерными нарушениями метаболизма, связанными с повышенным распадом белков, глюконеогенезом, перераспределением липидов. Однако интенсивность этих проявлений меньше, чем при первичном гиперкортизолизме, вследствие анаболических эффектов андрогенов, секреция которых также стимулируется избытком АКТГ. У женщин при этом в значительной степени проявляются вирилизм, гирсутизм, нарушение менструаций. Схемы соотношений в гипоталамо-гипофизарно-адреналовой оси в норме и при болезни Иценко-Кушинга представлены на рис. 5 и 6. Лечение болезни Иценко-Кушинга путем двусторонней адреналэктомии приводило к развитию синдрома Нельсона, который проявляется гиперпигментацией кожных покровов и слизистых оболочек и хронической недостаточностью надпочечников. Нарушение обратной связи приводит к разрастанию аденомы и дальнейшей гиперсекреции АКТГ, с чем связывают усиление его меланоцитстимулирующего эффекта.
Меланотропины α- и β-МСГ действуют на пигментные клетки (меланоциты). Вызывают распределение пигмента вдоль островков клеток, вследствие чего кожа темнеет. При недостатке гормона глыбки пигмента концентрируются в центре клетки – кожа светлеет. Гормон участвует в темновой адаптации глаза, повышает остроту зрения. Установлено также нейротропное действие МСГ: улучшение внимания, восприятия и кратковременной памяти. Кроме того, следует отметить антипиретические эффекты меланокортинов. Так, α-МСГ ослаблял лихорадку при центральном введении в дозах, которые не оказывали влияния на нормальную температуру тела. АКТГ1-24 и АКТГ1-39, содержащие такую же последовательность аминокислот, также ослабляли лихорадку. Эффект сохранялся у адреналэктомированных животных. Эти эффекты α-МСГ и его присутствие в мозге позволили предположить, что этот пептид может быть эндогенным модулятором лихорадки. В экспериментах на животных установлено, что во время лихорадки увеличивается его содержание в медиобазальном гипоталамусе и перегородке. Инактивация же пептида, образующегося в естественных условиях, ослабляла лихорадку, но не влияла на нормальную температуру тела. Антипиретические эффекты α-МСГ опосредуются меланокортиновыми рецепторами 4-го типа.
Липотропины Липотропин (β- и γ-) обладает липотропной активностью, а также некоторой кортикотропной и меланоцитстимулирующей активностью. Механизм липотропного действия: активация липазы жировой ткани → липолиз и освобождение жирных кислот. Кроме того, липотропины ускоряют обмен глюкозы в жировой ткани. Эндорфины Эндорфины относятся к группе опиоидных пептидов, рецепторы которых обнаружены, в первую очередь, в мозговых структурах, имеющих непосредственное отношение к проведению болевых импульсов и восприятию боли (околоводопроводное серое вещество среднего мозга, лимбическая система, спинной мозг). Эндорфины и другие опиоидные пептиды – энкефалины - обнаружены в различных отделах мозга.
Рис. 6. Изменения соотношения между компонентами гипоталамо-гипофизарно-адреналовой оси при болезни Иценко-Кушинга, вызванной аденомой аденогипофиза. Утолщенные линии обозначают усиление влияний и функции, пунктирные – ослабление влияний. Остальные обозначения как на рис. 5.
Биологические эффекты: 1. Выполнение функции медиаторов анальгетической (антиноцицептивной системы. 2. Влияние на эмоциональное состояние – формирование чувства удовлетворения, успокоения (поэтому эти пептиды называют пептидами вознаграждения), а также радостного, приподнятого настроения – эйфории. 3. Влияние на обучение и память. 4. Модулирующее действие на развитие иммунного ответа. 5. Участие в регуляции секреции пролактина, СТГ, ЛГ. 6. Стресс-лимитирующее действие.
Соматотропный гормон Соматотропный гормон (СТГ), или гормон роста (ГР) – пептидный гормон, содержащий 191 аминокислотный остаток. Особенности механизмов действия на ткани. На основании сопоставления метаболического действия СТГ in vivio и in vitro было сделано предположение об опосредованном участии его эффектов специальными медиаторами, образующимися в организме. В дальнейшем было установлено существование таких посредников, названных соматомединами. В настоящее время известно 4 соматомедина: А, А1, В и С. Они представляют собой полипептиды, синтезируемые в печени. Однако не исключается возможность их продукции в других тканях. Соматомедины (в первую очередь соматомедин С) вовлечены в регуляцию секреции СТГ по механизму отрицательной обратной связи. В торможении секреции гормона принимает участие соматостатин, продуцируемый в гипоталамусе. Эффекты СТГ В настоящее время эффекты СТГ разделяют на:
1) прямые: стимуляция синтеза соматомединов, в первую очередь, - соматомедина С (синоним – инсулиноподобный фактор роста-1 или ИФР-1) в печени и других тканях, стимуляция липолиза в жировой ткани и продукции глюкозы в печени;
2) непрямые: анаболическое действие и стимуляция роста. Эти эффекты опосредуются ИФР-1 и подавляются глюкокортикоидами. Схема регуляции выделения ГР представлена на рис. 7. Из рисунка видно, что секреция ГР стимулируется ГР-рилизинг гормоном (ГРРГ) и угнетается соматостатином. Отрицательная обратная связь на уровне гипофиза осуществляется соматомедином С (инсулиноподобный фактор роста-1). Этот фактор действует также на уровне гипоталамуса и стимулирует секрецию соматостатина. Кроме этого, на основе фармакологических данных установлено, что выделение ГРРГ стимулируется ацетилхолином, α-адренергическими и допаминергическими влияниями, но угнетается β-адренергическими. Секреция соматостатина (in vitro) стимулируется ацетилхолином и VIP и угнетается ГАМК. В естественных условиях выделение ГР модифицируется эндогенными ритмами сна, стрессом и физическими упражнениями.
Механизмы эффектов Метаболическое действие Белковый обмен: усиление синтеза белков. Механизмы: гормон создает условия для проникновения аминокислот через клеточную мембрану посредством повышения ее проницаемости, усиливает включение аминокислот в белки цитоплазмы клеток костной ткани, мышц, печени и почек, уменьшает активность протеолитических ферментов, стимулирует синтез рибонуклеиновой кислоты. Анаболическое действие СТГ проявляется в присутствии инсулина. Оно усиливается также малыми дозами глюкокортикоидов, но угнетается большими дозами.
Рис. 7. Регуляция секреции гормона роста. Условные обозначения как на рис. 5. Углеводный обмен. Анаболическое действие на углеводный обмен требует наличия инсулина. Поэтому при гиперпродукции СТГ сначала происходит активация, затем – истощение клеток и при длительном действии СТГ способствует развитию инсулинорезистентности и сахарного диабета. Действие на уровень глюкозы двоякое: сразу после введения СТГ увеличивает потребление глюкозы тканями и угнетается активность гексокиназы (уменьшается всасывание глюкозы в кишечнике), что приводит к гипогликемии, затем - гипергликемия. Механизм эффекта связывают с разной активностью N-концевой части молекулы (инсулиноподобное действие) и С-концевой (диабетогенная активность).
Механизмы развития гипергликемии при избытке СТГ: Ø СТГ → стимуляция α-клеток поджелудочной железы → усиление выработки глюкагона → активация гликогенолиза → гипергликемия; Ø СТГ → активация инсулиназы печени → разрушение инсулина → гипергликемия; Ø СТГ + глюкокортикоиды → торможение поглощения глюкозы тканями → гипергликемия. Жировой обмен: гормон повышает чувствительность адипоцитов к тоническим липолитическим влияниям, активность липаз и усиливает липолиз (жиромобилизующее действие). При избытке СТГ на фоне больших концентраций глюкокортикоидов усиливается липолиз в жировой ткани, на фоне высоких концентраций - замедляется. Считают, что влияние СТГ на жировой и белковый обмен не требует участия соматомединов. Лактогенное действие. Проявляется у человека и высших обезьян и обусловлено метаболическим влиянием на ткань молочных желез.
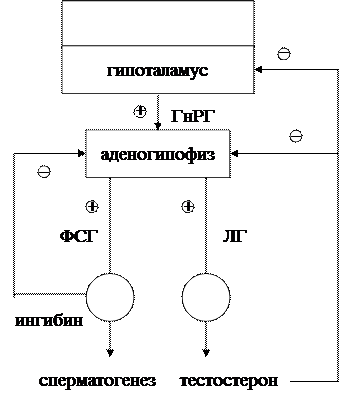
2. Ростовые эффекты. Существуют данные о различной локализации метаболической и ростовой активности в пределах молекулы гормона. Наиболее отчетливый эффект - стимуляция роста хряща. Механизмы: Ø митогенное действие (увеличение числа митозов в ткани); Ø усиление синтеза белка, включения тимидина в ДНК, синтеза РНК, образования хондроитин-сульфата. Эффекты опосредуются соматомединами, в первую очередь соматомедином С. Могут играть значительную роль в процессах регенерации тканей, заживления ран, в развитии компенсаторной гипертрофии после удаления одного из органов (например, почки). В последние годы появились данные о влиянии СТГ на рост лимфатических желез, лейкопоэз в костной ткани и функцию вилочковой железы. Гонадотропные гормоны Гонадотропины – ФСГ и ЛГ - по своей структуре являются гликопротеидами. ФСГ стимулирует у мужчин клетки Сертоли и сперматогенез. Под влиянием гормона эти клетки продуцируют белок, связывающий поступающие в семенную жидкость андрогены, и особый белок – "ингибин", который участвует в регуляции секреции ФСГ по механизму отрицательной обратной связи. ФСГ повышает чувствительность семенников к влиянию тестостерона. У женщин ФСГ стимулирует рост и созревание фолликулов. ЛГ у мужчин стимулирует интерстициальные клетки Лейдига, продуцирующие тестостерон. В связи с этим второе название ЛГ – "гормон, стимулирующий интерстициальные клетки". У женщин ЛГ активирует синтез и секрецию эстрогенов, овуляцию и образование желтого тела. Секреция ЛГ передней долей гипофиза контролируется отрицательной обратной связью: она обеспечивается тормозным действием половых гормонов на переднюю долю гипофиза и соответствующие нейроны гипоталамуса.
Пролактин Пролактин относится к белковым гормонам, пептидная цепь которого состоит из 198 аминокислотных остатков. Основные эффекты. 1. У самок млекопитающих возбуждает образование молока в молочных железах, действуя непосредственно на ферментативные системы железистых клеток. 2. Обладает лютеотропным действием – поддержание существования желтого тела и активация секреции прогестерона. 3. Тормозит секрецию гонадотропных гормонов. 4. У самцов млекопитающих стимулирует продукцию тестостерона, способствует развитию мужских половых органов, модулирует чувствительность семенников к действию гонадотропинов (ЛГ, ФСГ) и тестостерона. 5. Необходим для созревания легких у плода, способствует синтезу сурфактанта. 6. Обладает нейротропным действием – влияет на формирование родительских инстинктов. 7. Является лигандом для соответствующих рецепторов Т-лимфоцитов и, следовательно, может влиять на иммунные реакции. Установлена внегипофизарная продукция пролактина, в частности, фибробластами соединительной ткани, тимоцитами и лимфоцитами человека, клетками слизистой оболочки кишечника, эндометрия. Местно секретируемый пролактин может оказывать паракринное и аутокринное действие. В частности, имеется предположение о его иммуномодулирующем эффекте. Что касается пролактина, секретируемого децидуальными клетками эндометрия, то, согласно современной гипотезе, он предупреждает нарушение имплантации, подавляет сократительную деятельность матки при беременности, способствует синтезу сурфактанта у плода. В результате длительного повышения секреции пролактина (гиперпролактинемии) у женщин развивается синдром персистирующей аменореи и галактореи. Что касается патогенеза нарушений, то они могут иметь первичное гипоталамическое происхождение с ослаблением тонических дофаминергических влияний, которые в норме обеспечивают тормозный контроль секреции пролактина. Причиной первичных гипоталамических расстройств могут быть нейроинфекции, травмы черепа, в том числе перинатальные, с последующим повышением внутричерепного давления. Однако, как показали классические исследования, у многих больных выявляется первично гипофизарное заболевание (аденома-пролактинома) в результате мутации или избытка факторов роста. Вторичные (симптоматические) гиперпролактинемии могут возникать при избыточной секреции половых гормонов (синдром Штейна-Левенталя), терапии эстрогенами и препаратами, ухудшающими дофаминергическую передачу (резерпин), аутоиммунных заболеваниях (ревматоидный артрит, диффузный токсический зоб, системная красная волчанка и др.). Гиперпролактинемия угнетает гипоталамо-гипофизарно-адреналовую систему, вызывая развитие импотенции и бесплодие у мужчин, галакторею и аменорею - у женщин.
ГЛАВА 5 ПАТОФИЗИОЛОГИЯ ЭПИФИЗА Эпифиз (шишковидная или пинеальная железа)
Состоит из паренхиматозной ткани, содержащей пинеалоциты и глиальные клетки, и образуется как покрытый соединительной тканью вырост крыши III желудочка мозга. Характеризуется высокой проницаемостью гематоэнцефалического барьера. Активность эпифиза усиливается в темноте и ослабевает на свету и, таким образом, зависит от периодичности освещения. Тормозное влияние на железу реализуется через активность симпатической нервной системы. Эпифиз продуцирует в основном мелатонин, предшественником которого является серотонин, синтезируемый из триптофана. Синтез серотонина усиливается норадренергическими влияниями, а высвобождение из клеток – холинергическими. Однако, по мнению ряда исследователей, истинными гормонами эпифиза являются полипептиды, в частности пептид с антигонадотропной активностью и аргинин-вазотоцин.
ГЛАВА 6 Мужские половые гормоны К мужским половым гормонам относятся тестостерон, андростендион, дегидроандростерон. Основным гормоном является тестостерон: секретируется пульсативно (более 10 пиков за сутки). В крови связывается с транспортным глобулином. Схема взаимодействий в гипоталамо-гипофизарно-гонадной системе у мужчин представлена на рис. 8. Из рисунка видно, что секреция тестостерона стимулируется лютеинизирующим гормоном (ЛГ), в то время как созревание и рост тубулярных клеток активируются фолликулостимулирующим (ФСГ). Секреция тестостерона, в свою очередь, угнетает выделение ЛГ и ФСГ. Вероятно, что главная мишень для тестостерона – гипоталамус: он практически не влияет на эффективность действия ГнРГ (гонадотропин-рилизинг гормона) на гипофиз. Ингибин секретируется тубулярным эпителием и осуществляет прямое тормозное влияние на секрецию ФСГ. Секреция ГнРГ регулируется с участием аминергической системы мозга.
Физиологические эффекты 1. Формирование половых признаков. Если кастрация производится задолго до наступления половой зрелости, то наблюдается недоразвитие половых органов, а также вторичных половых признаков (строение скелета, мускулатуры, гортани, рост бороды и т.д.). После кастрации в зрелом возрасте некоторые вторичные половые признаки сохраняются (независимые половые признаки). 2. Андрогены влияют на развитие добавочных половых желез (предстательная железа, семенные пузырьки). Увеличивают в семенниках число рецепторов к ФСГ. 3. Влияние на скорость закрытия эпифизарных зон роста. При недостатке андрогенов в результате запаздывания окостенения эпифизарных хрящей увеличивается длина костей. 4. Анаболическое действие на обмен белков (в печени, почках, особенно в мышечной ткани). 5. Нейротропное действие: необходимо для формирования полового инстинкта, определяет агрессивность самцов.
Рис. 8. Схема взаимодействий в гипоталамо-гипофизарно-гонадной системе у мужчин. Условные обозначения как на рис. 5.
Основные формы патологии Гипогонадизм Первичный гипогонадизм развивается вследствие "железистых" причин снижения секреции андрогенов. В зависимости от этиологии и патогенеза выделяют врожденные и приобретенные формы синдрома. 1. Врожденные формы возникают при аплазии яичек, хромосомных нарушениях с кариотипами 47 XXY, 47 XYY, 48 XXYY, крипторхизме. Наиболее общим типом гипогонадизма у мужчин вследствие хромосомных нарушений является синдром Клайнфельтера, обусловленный присутствием одной или более лишней Х-хромосомы. Тяжесть клинических проявлений зависит от числа Х-хромосом. При классическом синдроме Клайнфельтера все клеточные линии имеют кариотип ХХY. Развитие характерных симптомов обусловлено недоразвитием яичек и снижением синтеза андрогенов. Кариотипу ХХY соответствуют евнухоидный внешний вид, гинекомастия (увеличение грудных желез), проявление женских вторичных половых признаков, включая отложение жировой ткани на бедрах и особенности оволосения. Крипторхизм – особый вид гипогонадизма, вызванный неопущением яичек в мошонку, когда они остаются в брюшной полости или в паховом канале, т.е. в условиях более высокой температуры. Результатом этого является недоразвитие яичек и проявления гипогонадизма. Крипторхизм может быть идиопатическим, но некоторые случаи ассоциированы со специфическим генетическим фактором, таким как трисомия по хромосоме 13. Имеются, однако, данные о том, что крипторхизм может развиваться как вторичное нарушение в результате недостаточной секреции лютеинизирующего гормона. 2. Приобретенные формы первичного гипогонадизма – результат прямого повреждения ткани железы при травмах, инфекции (туберкулез, сифилис, гонорея, эпидемический паротит), интоксикации (алкоголь, свинец), ионизирующей радиации, аллергических реакциях, кахексии, воспалении (орхит), гидроцеле, расширении вен семенного канатика, кастрации. Кастрация мужчин до наступления половой зрелости приводит к развитию евнухоидизма. Его проявления: чрезмерный рост костей в длину, недоразвитие и слабость мышц, высокий тембр голоса, развитие подкожной жировой клетчатки по женскому типу, женский тип оволосения, недоразвитие половых органов, отсутствие полового влечения и способности к половому акту. Кастрация взрослых мужчин сопровождается некоторой атрофией половых органов, обратным развитием вторичных половых признаков, снижением либидо и потенции.
Вторичный гипогонадизм возникает вследствие гипоталамо-гипофизарной недостаточности (недостаточная секреция гонадотропных гормонов, СТГ, ТТГ, тиреоидных гормонов). Примером вторичного гипогонадизма является адипозогенитальный синдром (синдром Фрёхликса) или гипоталамический евнухоидизм. В основе его – генетическая неспособность гипоталамуса секретировать нормальное количество гонадотропин-рилизинг гормона. Помимо признаков гипогонадизма, наблюдается ожирение.
Женские половые гормоны К женским половым гормонам относятся эстрадиол, эстриол, прогестерон. Основным эстрогенным гормоном является эстрадиол, менее активные – эстрон и эстриол. Все они – стероидные гормоны. В желтом теле яичников синтезируются прогестины (гестагены); основным гормоном в данной группе является прогестерон. Схема взаимодействий в гипоталамо-гипофизарно-гонадной системе у женщин иллюстрирована рис. 9. Схема показывает взаимодействие между невральным и гормональным контролем по механизму обратной связи. Развитие овариального фолликула находится под контролем ФСГ. Овуляция стимулируется ЛГ. Эстрогены имеют слож   Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования...  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|

 Курск – 2006
Курск – 2006




 Рис. 5. Схема соотношений в гипоталамо-гипофизарно-адреналовой оси в норме. Условные обозначения:
Рис. 5. Схема соотношений в гипоталамо-гипофизарно-адреналовой оси в норме. Условные обозначения:
 – активирующие влияния,
– активирующие влияния, – угнетающие влияния.
– угнетающие влияния.