|
|
Динамика условнорефлекторной деятельностиВ процессе жизнедеятельности животных и человека происходит постоянная смена репертуара приобретенных реакций. Одни условные рефлексы закрепляются, другие устраняются, одни рефлексы сменяют другие. Успех индивидуальной адаптации организма к разнообразным условиям его жизни, вея динамика условнорефлекторной деятельности зависят от сложных взаимоотношений между возбудительными и тормозными процессами в головном мозге. Тормозные процессы в условнорефлекторной деятельности. Заслуга открытия торможения в центральной нервной системе принадлежит И.М. Сеченову. Изучая закономерности условнорефлекторной деятельности мозга, И.П. Павлов установил, что существует два вида торможения — безусловное (внешнее) и условное (внутреннее). Внешнее (безусловное) торможение есть процесс экстренного ослабления или прекращения отдельных поведенческих реакций при действии раздражителей, поступающих из внешней или внутренней среды. Это торможение возникает без какой-либо предварительной выработки, т. е. является врожденным свойством нервной системы. Внешнее торможение различных условнорефлекторных реакций, возникающее при действии посторонних стимулов, часто встречается в повседневной жизни человека и животных. Причиной могут быть различные условнорефлекторные реакции, а также различные безусловные рефлексы (например, ориентировочный рефлекс, оборонительная реакция — испуг, страх). Однако в большинстве случаев посторонние стимулы вызывают ориентировочную реакцию, что соответственно приводит к прекращению текущей поведенческой деятельности организма. При длительном или повторном действии стимула или его узнавании происходит угашение ориентировочного рефлекса, что выражается в нивелировании тормозного состояния и в восстановлении исходного уровня условнорефлекторной деятельности (гаснущий тормоз). Другой разновидностью врожденного тормозного процесса является так называемое запредельное торможение. Оно развивается при длительном нервном возбуждении организма. Экспериментально запредельное торможение развивается при воздействиях сильных раздражителей или нескольких несильных, суммарный эффект которых при одновременном и длительном применении превышает предел работоспособности нервных клеток мозга. Это торможение называют также охранительным, полагая, что оно предохраняет нейроны от истощения. Временно выключая активность нервных клеток, оно создает им условия для восстановления нормальной возбудимости и работоспособности. Несомненно, явление запредельного торможения наблюдается тогда, когда сильное волнение вызывает у некоторых людей заторможенное состояние. Условное (внутреннее) торможение является приобретенным и проявляется в форме задержки, угашения, устранения условных реакций. Условное торможение является активным процессом в нервной системе, развивающимся, как и условное возбуждение, в результате выработки. Такое внутреннее торможение выполняет важнейшие функции перестройки в условнорефлекторной деятельности. Выделяют четыре вида внутреннего торможения: угасательное, дифференцировочное, запаздывательное и условный тормоз. Общим для всех видов внутреннего торможения является их развитие на базе предварительно выработанного условного рефлекса. Угасательное торможение (угашение условного рефлекса) возникает после отмены подкрепления условного стимула. Условный стимул теряет свое первоначальное значение. Скорость угашения условной связи находится в обратной зависимости от интенсивности условного стимула и силы, биологической значимости подкрепления. Усиление внутреннего (угасательного) торможения может привести к угнетению не только угашаемого рефлекса, но и других, прежде всего близких, однородных условных рефлексов (вторичное угашение). Торможение, лежащее в основе угашения, является важной биологической функцией, так как оно способствует устранению условных рефлексов, ставших уже бесполезными, ненужными. Механизм угасательного торможения, таким образом, снижает активность «ненужных» функциональных связей в мозге. В изменившихся условиях существования организма будут вырабатываться и новые адаптивные реакции. Дифференцировочное торможение является важным механизмом в деятельности мозга, способствующим различению сигналов. Тонкое различение сигнального раздражителя происходит в результате неподкрепления посторонних стимулов, близких по своим параметрам условному сигналу. Развитие этого вида торможения идет постепенно (в три этапа). Когда на фоне выработанного условного рефлекса применяется дифференцировочный (новый) раздражитель, то часто он тоже вызывает условный ответ, хотя и с меньшим эффектом, чем условный подкрепляемый стимул. Это связано с тем, что дифференцировочный раздражитель, как всякий новый стимул, вызывает ориентировочный рефлекс, который обусловливает внешнее торможение условного ответа (I этап). По мере дальнейшего попеременного применения условного и дифференцировочного раздражителей ориентировочная реакция на сходный раздражитель исчезает, вследствие чего условные реакции на оба стимула выравниваются (II этап). Затем на неподкрепляемый дифференцировочный раздражитель условная реакция угасает (III этап). Дифференцировка (различение) стимулов вырабатывается тем быстрее и легче, чем отдаленнее сходство между раздражителями. При очень близких сигналах дифференцировка может совсем не образоваться в результате первичного дефицита информации, воспринимаемой организмом. Запаздывательное торможение образуется при выработке запаздывающих и следовых условных рефлексов, когда условный сигнал значительно опережает подкрепление. С помощью этого вида внутреннего торможения рефлекторная реакция может быть приурочена к моменту предъявления подкрепления. Развитие этого торможения происходит во время изолированного (опережающего) действия условного стимула. Постепенно условная реакция смещается во времени ближе к моменту появления подкрепления. Запаздывательное торможение, т. е. способность нервной системы к «экономному» режиму работы, проявляется различно у возбудимых и инертных животных. У возбудимых животных запаздывательное торможение развивается труднее и медленнее. Условный тормоз развивается в том случае, если условный сигнал в сочетании с каким-либо дополнительным агентом не подкрепляется, а изолированное действие условного стимула подкрепляется. Вскоре условный стимул в комбинации с дополнительным агентом перестает вызывать условную реакцию благодаря развитию условного тормоза. Биологическое значение условного тормоза состоит в уточнении сходства и различия сигналов. Рассматривая варианты внутреннего торможения, П.К. Купалов считал, что запаздывательное торможение и условный тормоз есть, по сути, частные случаи дифференцировочного торможения. В ответ на многократные или монотонные стимулы непременно развивается внутреннее торможение. Если такая стимуляция продолжается, то наступает сон. Переходный период между бодрствованием и сном назван гипнотическим состоянием. И.П. Павлов разделил гипнотическое состояние на три фазы в зависимости от размеров области коры полушарий, охваченной торможением, и соответствующей реактивности различных мозговых центров в процессе реализации условных рефлексов. Первая из этих фаз называется уравнительной. В это время сильные и слабые стимулы вызывают одинаковые условные ответы. Парадоксальная фаза характеризуется более глубоким сном. В этой фазе слабые раздражители вызывают более интенсивный ответ, чем сильные. Ультрапарадоксальная фаза означает еще более глубокий сон, когда ответ вызывают только слабые стимулы, а сильные приводят к еще большему распространению торможения. За этими тремя фазами следует глубокий сон. И.П. Павлов считал, что гипнотическое состояние является частичным сном. Однако оказалось, что эта проблема не так проста. Было установлено, что в гипнозе волны электроэнцефалограммы (ЭЭГ) сходны с ЭЭГ бодрствования, а не сна. Выяснилось, что одни люди более подвержены гипнозу, чем другие, а некоторые вовсе не поддаются ему (что говорит о различном соотношении процессов возбуждения и торможения у разных людей). В последние годы получили распространение теории, согласно которым гипнотическое состояние нельзя приравнивать ни к сну, ни к бодрствованию. Свойства основных нервных процессов. Неразрывное единство двух противоположных нервных процессов — возбуждения и торможения — составляет фундаментальный механизм всей условнорефлекторной деятельности. Оба нервных процесса участвуют в формировании условного рефлекса, сложным образом взаимодействуя между собой. Факт распространения (иррадиации) возбуждения по нервной системе давно известен физиологам и клиницистам. Н.Е. Введенский показал, что иррадиация — это основное свойство нервного процесса и что возбуждение, возникшее в одном центре, может распространяться и на другие центры. Так, при развитии эпилептического припадка возбуждение, возникшее в эпилептоидном очаге, постепенно охватывает вею двигательную кору. Эффекторная генерализация проявляется вначале в виде усиления реакций отдельных мышц, а затем в реакцию вовлекается вся скелетная мускулатура и ряд вегетативных изменений. Если возбуждение распространяется и на зрительную область коры, то возникают зрительные галлюцинации. В клинике известны также случаи иррадиации болевых ощущений. Движение нервных процессов зависит от силы. Как правило, иррадиация (распространение) нервных процессов проявляется при слабой интенсивности возбуждения и торможения, т. е. на стадии слабых, неупроченных временных связей. При достаточной силе нервных процессов они концентрируются, а при чрезмерной силе вновь начинается процесс иррадиации. Дальность распространения нервных процессов от исходного пункта также зависит от их силы: чем интенсивнее нервный процесс в исходном пункте, тем дальше он распространяется и тем сильнее его воздействие на соседние участки мозга. Процесс формирования классического условного рефлекса проходит три основные стадии. Наиболее ранний период выработки временной связи, стадия прегенерализации, характеризуется значительными изменениями фоновой ритмики электрических потенциалов различных областей мозга, но при этом отсутствуют условные поведенческие реакции. Для этой стадии характерна выраженная концентрация возбуждения, главным образом в проекционных зонах коры условного и безусловного раздражителей. После этой кратковременной фазы концентрации возбуждения следует стадия генерализации условного рефлекса, в основе которой лежит процесс «диффузного» распространения (иррадиации) возбуждения. Во время стадии генерализации условные реакции возникают на сигнальный и другие раздражители (явление афферентной генерализации), а также в интервалах между предъявлениями условного стимула (межсигнальные реакции). Различные биоэлектрические сдвиги (блокада альфа-ритма, десинхронизация, тета-ритм и вызванные потенциалы) широко распространены по коре и подкорковым структурам. Далее по мере подкрепления условного стимула межсигнальные реакции угасают и условный ответ возникает только на сигнальный раздражитель. Это — стадия специализации. На этой фазе выработки временной связи изменения биотоков более ограничены и приурочены в основном к действию условного стимула. Этот процесс обеспечивает дифференцировку, тонкое различение стимулов, специализацию условнорефлекторного навыка. В случае выработки инструментального рефлекса стадия генерализации четко проявляется в форме «диффузной» двигательной активности. Явление эфферентной генерализации проявляется в целом наборе двигательных реакций на действие одного и того же условного стимула. На стадии специализации временной связи условный стимул запускает узкоспециализированную двигательную активность. Развитие таких процессов можно наблюдать у человека при выработке различных профессиональных навыков (по типу инструментального научения), например, работа на конвейере, печатной машинке, овладение детьми умением письма и др. Детальное исследование динамических изменений электрической активности мозга проведено в лаборатории М.Н. Ливанова с помощью ЭВМ и многоточечного отведения биопотенциалов различных областей мозга. Была установлена функциональная связь синхронных отношений биопотенциалов мозга человека при реализации двигательных реакций, а также при выполнении умственной деятельности (рис. 16). Исследование дистантной синхронизации (сходство биопотенциалов разных отделов мозга) на животных проведено на модели выработки условного рефлекса (например, сочетание света и болевого раздражения конечности кролика). Было показано, что в период генерализации условного рефлекса наблюдается синхронизация биоэлектрической активности во многих Рис. 16. Временная корреляция биопотенциалов точек коры головного мозга человека в процессе умственной деятельности (по М.Н. Ливанову, Т.П. Хризман, 1976). А — состояние покоя, Б — через 15 с от начала решения задачи. Кружками обозначены электроды, заштрихованные кружки — точки корреляции
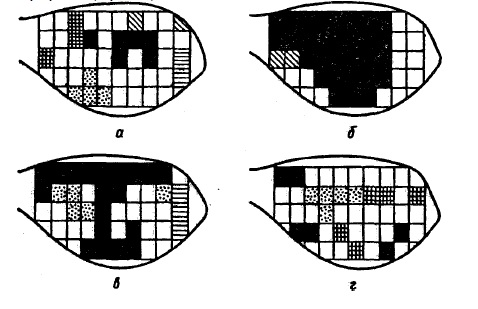
участках коры и подкорковых образованиях (рис. 17). В процессе специализации сфера распространения этих потенциалов значительно сужена и возрастает условнорефлекторный ответ. Необходимым условием образования функциональной связи между корковыми центрами ассоциируемых раздражителей является синхронизация биопотенциалов (в диапазоне тета-ритма). Иначе говоря, условная реакция животного при сочетании света и болевого раздражения конечности возникала при высокой степени сходства биопотенциалов зрительной и двигательной областей коры. Рис. 17. Динамика синхронизации волн электрокортикограммы кролика в процессе выработки условного рефлекса (по M Л. Ливанову, 1972). Квадраты — пункты отведения, одинаково заполнены участки, дающие синхронизацию волн от 70 до 100% времени измерения; а — исходный фон, б — после 12 сочетаний условного и безусловного стимулов, в — после 60 сочетаний (выработан условный ответ), г — после 90 сочетаний (условный рефлекс упрочен)
Из вышеизложенного следует, что распространение нервных процессов и их взаимное влияние друг на друга тесно связаны между собой. Изменения возбудимости в различных участках мозга происходят «волнообразно», или, по выражению И.П. Павлова, в коре наблюдается «функциональная мозаика». Конфигурация ансамбля активно действующих зон (пространственно распределенных локусов) не останется застывшей, она меняет свою пространственно-временную конфигурацию. Такую функциональную мозаику очагов возбуждения удается наблюдать по изменениям электрографических показателей активности при многоканальном отведении с использованием ЭВМ и регистрацией на экране осциллографа или телевизора или с помощью тепловизора.   Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|