|
|
ГРИБЫ И ГРИБОПОДОБНЫЕ ОРГАНИЗМЫСтр 1 из 4Следующая ⇒ ГРИБЫ По определению американского ботаника Р. Уитгейкера, к грибам следует относить гетеротрофные эукариотнме организмы с осмотрофным питанием. С особенностями питания связаны характерные черты строения, образа жизни и химических свойств грибов. 1. Наиболее распространенной формой вегетативного тела грибов (таллома) является мицелий, или грибница. Это многократно ветвящиеся нити (гифы), которые пронизьивают субстрат, будь то почва, ткань растения или навозная куча. Такое строение оптимально для всасывания из субстрата питательньтх веществ всем телом. 2. Большинство оргашческих веществ в субстрате находится в форме биополимеров — полисахаридов, белков и крупных молекул других веществ, неспособных проникать через клеточную мембрану. Гифы грибов, вьщеляя в окружающую среду активные ферменты деполимеразы, разрушают полимерьи до способных транспортироваться в клетку моно- или олигомеров. 3. Клетки грибов развивают огромное, несопоставимое с клетками других эукариотных организмов тургорное давление, позволяющее им, как насосу, всасывать из окружающей среды растворенньие в воде питателыые вещества. 4. Поскольку все вегетативное тело погружено в субстрат, у многих грибов возникают проблемы с распространением спор. Поэтому большинство грибов формируют споры на специальных спороносцах, поднимающихся над субстратом. То, что в просторечье называют «грибами», является их спороносящими органами. Такие морфологические и эколого-трофические особенности присущи большой группе организмов, которые в предыдущем издании учебника рассматривались в качестве классов единого отдела «Грибы». Однако уже в то время было известно, что некоторые грибы имеют кардинальные отличия в ультраструктуре жгутиков и митохоцдрий, химическом составе и других свойствах от остальных грибов. По перечисленным признакам они имели больше сходства с гетероконтными (разножгутиковыми) желто-буроокрашенными водорослями, чем с другими грибами. Эти различия были подтверждены молекулярньими филогениями. Показано, что большинство грибов имеет монофилетическое (от одного корня) происхождение и формирует самостоятельное царство Еumycota. Классы Нурochytriomycetes, Оomycetes и сетчатые слизевики, относимые ранее к миксомицетам, имеют общее происхождение с отделом Осhrophyta и образуют вместе с ним самостоятельное царство Stramenopila. долтое время господствовало представление, что оомицеты — ветвь гетероконтных водорослей (на основании морфологических аналогий их относили к желтозеленым), потерявшая хлоропласты. Многие соображения, в том числе и те, которые основаны на данных молекулярной филогении, показывают, что эти организмы составляют ветвь страменогшл, отклонившуюся до того, как зоофлагеллятный предок приобрел в качестве хлоропласта одноклеточную красную водоросль. В связи с вышесказанным многие микологи вообще не рекомендуют называть оомицеты и родственные им группы «грибами», предлагая такие названия, как «псевдогрибы», «грибоподобные организмы» и т.п. В предлагаемом учебнике они рассматряваются не среди грибов, как в предьщущем издании, а отдельно, предваряя разбор филогенетически единого царства Еumycota. Что касается «истинных грибов», то изменения их системы, произошедшие в последние годы, подробно рассмотрены в соответствующем разделе. Здесь лишь сделаем два замечания: 1) поскольку статус грибов поднят с отдела до царства, отдельные классы в данном пособии рассматриваются как отделы; 2) с позиций филогении класс Deuteromycetes (несовершенные грибы), в который объединяли сумчатые (главным образом) грибы, вторично утратившие половое спороношение, упраздняется, ибо входящие в него организмы полифилетичны (произошли от разных предков). Рассмотрение дейтеромицетов как отдельной группы в данном руководстве, хотя и с оговорками, вызвано многообразием этих грибов и той важной ролью, которую они играют в природе и хозяйственной деятельности. Лишайник представляет собой симбиотический организм, в котором клетки водоросли оллетены гифами гриба. Такой симбиоз обусловил специфические особенности морфологии, физиологии, биохимии и образа жизни лишайников, представляющих собой четко отграниченную в морфологическом и физиологическом отношении группу, которую изучают специалисты именно по этой группе — лихенологи. Однако лишайники — полифилетическая группа, ибо их тело образовано грибами и водорослями из разных отделов и классов. Тело лишайника образовано грибным талломом, а водоросли находятся обычно внутри него, поэтому в современных системах ляшайнихи разбросаны по тем группам грибов, которые входят в их состав. В данном учебнике лишайники рассматриваются отдельно по высказанным выше соображениям относительно своеобразия их строения и образа жизни. Миксомицеты, или слизевики, имеют одноклеточный амёбоидный или многоклеточный ллазмоидный таллом, не покрытый плотной оболочкой и способный к амёбообразным движениям. Для них характерно миксотрофное (смешанное) питание — путем всасывания растворенных питателъных веществ (как грибы) и заглатывании мелких бактериальных и дрожжевых клеток в пиноцитозный пузырек (как животные). На этом основании знаменитый немецкий ботаник и миколог А. де Вари назвал их Мусеогоа (грибоживотные). Согласно современным филогениям, большинство миксомицетов образует самостоятельное царство, примыкающее к амёбам, но есть группа (акразиевые), объединенная в одно царство с эвгленами. Итак, мы рассмотрели с разных позиций особенности тех групп организмов, которые включены в «Курс альгологии и микологии». Как видно, эти организмы выходят за рамки водорослей и грибов в узком понимании этих терминов. Они образуют большое число самостоятельно эволюционирующих групп (царств), причем у разных исследователей это число неодинаково. В табл. 1 приведены те царства, в которые входят рассматриваемые в учебнике организмы. В основу таблицы положена система, рассматриваемая в книгах О.Г. Кусакина и А.Л. Дроздова «Филема органического мира», с рядом модификаций, касающихся главным образом гругшы хромофитовых (в системе Кусакина и Дроздова — гетероконтных), которые, исходя из предложенного выше принципа брать за основу классификации хозяйскую, а не эндосимбиотическую клетку, разделены на три царства.
Таблица отчетливо показывает, что водоросли и грибы в широком смысле слова — понятия не филогенетические, ибо могут находиться не только в разных, но и в общих (одних и тех же) филах, которые могут объединять практически все эколого-трофкческие группы организмов — растения (водоросли), животные (простейшие) и грибы. Это, с одной стороны, свидетельствует о единстве живой природы, а с другой — усложняет принципы изложения материала в учебнике, где в целях оптимального восприятия нужны четкие разделительные признаки, которых, к сожалению, в природе нет. Организмы, традиционно называемые «низшими растениями», являются важной составной частью биоты и неразрывно соединены многочисленными связями с другими организмами. Их изучение имеет огромное значение для многих разделов теоретической и прикладной биологии, важнейшие из которых следующие. • Как было сказано, эти организмы входят в состав многих независимых эво- люционных линий, различающихся не только происхождением, но и ультра- структурой, обменом веществ, образом жизни. «Низшие растения» — это полигон, на котором природа методом «проб и ошибок» создавала оптимальные конструкции, оказавшиеся способными стать доминирующими на суше и в море. В связи с этим их сравнительное изучение (молекулярно-биологическое, цитологическое, морфологическое, биохимическое) имеет огромное значение для понимания ранних и средних этапов эволюции биоты. • Изучение «низших растений» очень важно ддя решения одной из централь- ных задач современной биологии — «инвентаризации» биологического разнообразия. Согласно определению, принятому Международной конвенцией по биоразнообразию в Рио-де-Жанейро, «биологическое разнообразие означает вариабельность живых организмов из всех источников, включая среди прочего наземные, морские и иные водные экосистемы и экологические комплексы, частью которых они являются; это понятие включает в себя разнообразие в рамках вида, между видами и разнообразие экосистем». Если инвентаризация высших эукариот (высших растений и позвоночных животных) близка к завершению, то оценка разнообразия ряда групп низших организмов находится лишь на начальной стадии. Предполагается, что отписана только небольшая часть обитающих в морях грибов и «псевдогрибов», диатомовых и галофитовых водорослей. Огромное число неописанных видов грибов обитает в тропических лесах на листьях растений, существует в качестве паразитов и симбионтов насекомых и других беспозвоночных животных. Кроме того, в последние годы было установлено экспериментальными методами, что многие грибы, описанные как один вид на основании морфологического сходства, на самом деле включают ножество (до десяти и более) биологических нескрещивающихся видов-двойников, различающим многими физиологическтгми параметрами. • «Низшие растения» вносят огромный вклад в глобальную экологию, в част- ности в миграцию углерода. Морские водоросли — важнейший продуцент органического вещества в океане, на запасах которого существует вся пищевая лмрамида вплоть до крупных рыб и китообразных. С учетом вклада в фотосинтез пикопланктона (мелких одноклеточных цианобактерий и эукариотных водорослей) продукция связанного углерода в Мировом океане превышает продуктивность наземных растений. Мелкие одноклеточные морские водоросли, покрытые известковыми пластинками, — гаптофитовые, в последние 150 млн лет вносили главный вклад в связывание углерода углекислого газа кальцием и образование осадков известняка, покрывающих половину поверхности дна океана (треть всей поверхности Земли). Наземные грибы, наоборот, как важнейшие редуценты разлагают органические полимеры и освобождают из них углерод вплоть до углекислого газа. Огромные запасы связанного углерода накапливаются в древесине в виде очень стойких полимеров — целлюлозы и лигнина. Грибы (особен- но трутовые) — единственные организмы на Земле, обладающие ферментами, способными разрушить лигноцеллюлозный комплекс. Не будь их, лес был бы до макушек деревьев покрыт мертвыми ветками. Не менее важную роль играют грибы в мигращлт органических веществ в почве и сщцапии почненного гумуса. Через тело грибов проходит 1/2 запасенного органического углерода нашей планеты. Многие «низшие растения», особенно грибы, наносят огромный урон народному хозяйству и угрожают здоровью населения. Они вызывают наиболее разрушительные эпидемии растений. Ржавчина хлебных злаков, фитофтороз картофеля, ожог листьев риса, корневые гняля и другие болезни растений часто приводили и приводят к гибели урожая на огромных площадях. Поскольку многие грибы образуют токсические метаболиты, зараженная ими пища становится причиной массовых отравлений людей и скота. Дерматофиты, разлагающие белок кератин, который входит в состав покровных тканей (кожи, ногтей, волос), вызывают поверхностные микозы у диких и домашних животных и людей. В последние годы участились случаи гораздо более опасных — внутренних — глубоких микозов человека, часто приводящих к смертельному исходу. Наличие у грибов разнообразных ферментов позволило им освоить новые техногенные субстраты: заменители кожи, полимерьт, кяноггленки, стекло, бумагу, строительньте материалы. Поврежцение промышленных изделий наносит огромный материальный ущерб, а при разрушении грибами произведений искусства - картин, древних рукописей, исторических зданий — потери вообще невосполнимы. Мероприятия по защите всех этих разнообразных живых и техногенных объектов невозможны без детального изучения вредных грибов, их физиологяи и молекулярных основ взаимоотношений паразитов с их хозяевами. • Благодаря продукции разнообразных биологически активных соединений грибы и водорослм с древних времен являются обьекгами пристальною внимания биотехнологии. Так, первым промышленным микроорганизмом, используемым человечеством, были грибы — дрожжи, давшие людям важнейшие продукты — хлеб и вино. Антибиотики и другие лекарственные препараты, ферменты, витамины, органические кислоты, пищевые добавки, съедобные грибы и морские водоросли — вот далеко не полный перечень их биотехяологяческого использования. В связи с этим огромную роль играет практически только начатое изучение биоресурсов грибов и водорослей, т.е. генетяческях ресурсов организмов и популяций, имеющих фактическую или потенциальную ценность для человечества. Можно ли было предположить еще 20 лет назад, что многие древоразрушающие грибы окажутся источником замечательных лекарственных препаратов - противораковых, иммуностимулирующих и др.? • Будучи простейшими эукариотами, «низшие растения» стали излюбленной моделью экспериментальной биология. Клеточный миксомицет Diсtyostellium и зеленая водоросль Асеtabulariа— замечательные объекты для изучения биологии развития; дрожжи— первый эукариотный организм, у которого секвенирован весь геном и сделано много замечательных открытий, оказавших влияние на развитие молекулярной и клеточной биологии у всех организмов, вплоть до человека; зеленая водоросль, Chlamydomonas широко используется в работах по изучению молекулярной биологии хлоропластов; исследования генетической рекомбинации у сумчатых грибов заложяли основы для создания молекулярной теории рекомбинации. Но для изучения молекулярной биологии, генетика и биохимии этих и других объектов надо хорошо знать их биологию, а также уметь выбрать из разнообразия видов наиболее адекватный для решения новой экспериментальной задачи. Подходы к такой деятельности дает предлагаемый читателю «Курс альгологии и микологии». ОБЩАЯ ХАРАКТЕРИСТИКА Определение. С эколого-трофической точки зрения к грибам относят гетеротрофные эукариоты с исключительно осмотрофным типом питания, вегетативное тело которых обычно погружено в субстрат, а на поверхность субстрата выдвигаются спороносные органы. У многих грибов (макромицетов) они крупные и хорошо заметные — это, например, поднимающиеся над почвой плодовые тела шляпочных грибов или вырастающие на дереве трутовики. Другие грибы (микромицеты) имеют мелкие органы спороношения, строение которых можно рассмотреть только под микроскопом. При массовом развития они образуют цветные налеты в виде плесеней на различных субстратах. Филогенетические построения показывают, что экоморфа «грибы» не является однородной монофилетической группой, а разделяется на две филы (царства). Большая часть, названная «истинными грибами» (эумицеты), монофилетична и составляет царство Грибы (Fungi), которое разделяют на четыре отдела — Chytridiomicota, Zygomycota и два отдела — сумчатые (аскомицеты) и базидиальные грибы из надотдела Dikaryomycota. Меньшая часть, названная «грибоподобными организмами» (псевдомицеты), входит наряду с некоторыми водорослями в состав царства галiепорi1а, группируясь в два отдела — Оomycota (оомицеты) и Labyrinthulomycota (сетчатые слизевики). Грибной таллом Морфологическая классификация позволяет выделять три типа талломов, присущие большинству грибов: амёбоидный (плазмодиальный), мицелиальный и дрожжеподобный. Амёбоидный таллом. Представляет собой лишенную клеточной стенки одноядерную клетку. Кроме цитоплазматической мембраны клетка защищена лишь уплотненным периферическим белковым слоем — перипластом. Многоядерный таллом такого типа называют плазмодиальным. Для закрепления в субстрате и питания таллом, находящийся на поверхности (экстраматрикальный) или внутри (интраматрикальный) субстрата, может образовывать корнеподобные выросты — ризомицелий. Если ризомицелий отсутствует или лишен ядер, то таллом представлен одной амёбоидной клеткой и называется моноцентрическим. У некоторых грибов ризомицелий удлинен, в него переходят ядра, и он образует в некоторых местах расширения, образуя другие амёбоидной клетки с ядрами (полицентрический таллом).
У примитивных видов моноцентрический таллом при переходе к спороношению целиком превращается в спорангий (холокарпический таллом). У грибов с эукарпическим талломом из амёбоидной клетки формируется несколько спорангиев или они образуются в нескольких клетках полицентрического таллома. Мицелиальный таллом. Это покрытые клеточной стенкой разветвленные нити, разделившие внутренними перегородками на фрагменты (клеточный мицелий) или не фрагментированные (неклеточный мицелий). Перегородки образуются врастанием клеточных покровов (клеточной стенки и мембраны) от периферии к центру (подобно диафрагме микроскопа или оптического фотоаппарата). Они обычно не замкнуты, в середине остается просвет, через который из одной клетки в другую могут мигрировать питательные вещества и некоторые органеллы, включая ядра. В клетках многих сумчатых грибов находятся включения углеводной или белковой природы (пробки, тельца Воронина), которые могут закупоривать отверстие в септе и останавливать межклеточные обмены. Как правило, пора одна, но встречаются септы, перфорированные многими порами наподобие сита. Септы с простой порой могут постепенно утончаться к центру или иметь одинаковую толщину. Долипоровая септа, характерная для базидиомицетов, имеет трубчатое расширение у поры и прикрыта перфорированным мембранным колпачком — парентосомой (рис. 177). Через такую пору миграция клеточных органелл невозможна, хотя на отдельных стадиях жизненного цикла происходит растворение парентосом и появляется возможность межклеточных обменов органеллами.
Гифы имеют верхушечный рост, причем растет не только верхушка первичной гифы, но и веточки, поэтому в зрелом мицелии гифы настолько густо переплетены, что бывает невозможно определить, где первичная гифа, а где веточки. Картину усложняют многочисленные анастомозы (слияния) боковых веточек друг с другом (рис. 178). Анастомозы придают прочность мицелиальной структуре. Диаметр гиф обычно не бывает меньше 1 мкм и больше 30 мкм (в среднем около 10 мкм). Тем не менее у некоторых представителей рода Аchlya гифы могут достигать ширины 1000 мкм (т.е. 1 мм!). Гифы, составляющие мицелий, могут быть примерно одинакового диаметра, но могут сильно различаться (как стволы и ветви деревьев). Именно из мицелия (за редчайшими исключениями) формируются многочисленные структуры грибов, в том числе и плодовые тела, в обиходе называемые «грибами». Видоизменения мицелия обусловлены адаптациями к условиям жизни, ибо мицелий несет не только функции, обеспечивающие питание, но и выживание в окружающем мире. Все видоизменения можно свести к обеспечению двух функций — вегетативной и пропагативной. Вторая функция будет рассмотрена в последующих разделах, а здесь остановимся на первой. Вегетативная функция может быть разделена на обеспечение выживания (покоящиеся структуры), поддержание целостности мицелия в пространстве (проводящие и поддерживающие структуры), заражение и питание в теле хозяина (инфекционные структуры). Конечно, такое деление, как и всякая другая биологическая классификация, условно, ибо многие грибные структуры полифункциональны. Например, зиготические (т.е. пропагативные, а не вегетативные) структуры многих грибов покрыт толстыми оболочками и несут функцию выживания, а вегетативные хламидоспоры наряду с функцией выживания обеспечивают размножение. Среди покоящихся структур рассмотрим склероции и хламидоспоры. Склероции представляют собой тесное переплетение гиф (ложная ткань — плектенхима), образующее комочек размером от нескольких десятков микрометров до нескольких сантиметров. По анатомическому строению склероции разделяют на истинные и ложные. У первых на поперечном разрезе можно увидеть два слоя: узкий — коровый и широкий — сердцевидный (рис. 179).
При образовании хламидоспор отдельные клетки мицелия покрываются толстыми, часто меланизированными оболочками и обособляются. Хламидоспоры, одиночные или соединенные в цепочки, могут образоваться интеркалярно (из внутренних клеток мицелия) и терминально (из конечных клеток). Они могут быть различными по форме и размерам. • Проводящие в поддерживающие структуры характерны для древоразрушающих базидиомицетов, мицелий которых часто распространяется на очень большие расстояния вдоль дерева (иногда на десятки метров). Для обеспечения единства таллома на таком большом протяжении отдельные гифы срастаются боковыми сторонами, образуя пучки — синнемы. Крупные, прочные, видимые невооруженным глазом синнемы называют тяжами или шнурами. Центральные гифы в таких шнурах могуг функционировать как проводящая ткань, обеспечивая транспорт воды и питательных веществ. У осеннего опенка (Armillaria mellea) оболочки наружных клеток разветвленных тяжей меланизированы, вследствие чего они становятся очень прочными и темноокрашенными (похожими на проволоку). Такие «гибриды» тяжей и склероциев называют ризоморфами. Они способны распространяться не только по зараженному стволу дерева, но и в почве и заражать корни расположенных вблизи здоровых деревьев. • Инфекционные структуры предназначены для заражения растений или животных и питания за их счет. Паразиты растений часто образуют на поверхности расширение кончика гифы апрессорий, который плотно прилегает к кутикуле за счет выделяемых наружу гидрофобных белков гидрофобинов, служащих «вакуумной смазкой». На участке оболочки апрессория, не соприкасающемся с кутикулой, откладывается меланин, а в растение из апрессория выделяются гидролитические ферменты, разрушающие кутикулу и клеточную стенку растения. Под действием высокого турторного давления и ферментов в оболочке растения образуется отверстие, через которое прорастает внутрь инфекционная гифа, преобразующаяся в питательный орган — гаусторий. При росте гаустория плазмалемма хозяйской клетки не разрушается, а вдавливается (инвагинируется), вследствие чего зараженная клетка не погибает, а некоторое время снабжает гаусторий питательными веществами и обеспечивает внутритканевый (межклеточный) рост гиф паразита. По форме гаустории бывают простыми и разветвленными (рис. 180).
МКГ — материнская клетка гаустория (инфекционная гифа гриба); ЭМ — экстрагаусториальная мембрана (продолжение плазмалеммы растительной клетки, окружающее гаусторий); ЭГМ — эксграгаусториальный матрикс (пространство между ЭМ и клеточной стенкой гаустория)
Интересны по своему строению ловчие гифы хищных грибов, питающихся мелкими почвенными и водными нематодами, амёбами, ракообразными и другими беспозвоночными животными. Ловчие гифы могут представлять собой различного рода кольца, петли, сети, клейкие нити, стреляющие «гарпуны», которые образуются в ответ на присутствие определенных веществ потенциальной жертвы и сжимаются или выстреливают при механическом соприкосновении с ней. • Ложные ткани. Настоящие ткани, возникающие в результате деления клеток в разных направлениях, образуют немногие грибы, в частности лабульбениевые аскомицеты. Однако мицелий может формировать ложные ткани (плектенхимы), из которых, в частности, построены плодовые тела. На поверхности ризоморф, шляпок плодовых тел, склероциев образуется кроющая «ткань»; у некоторых трутовых грибов сильно развиты механические элементы, а млечные ходы некоторых видов шляпочных грибов, таких, как виды рода Lactarius (рыжики, грузди, волнушки), можно рассматривать как подобие проводящих тканей. В особенностях морфогенеза сложно устроенных плодовых тел различных грибов — макромицетов — также большую роль играют гидрофобины, обеспечивающие слипание соседних гиф. Дрожжеподобный почкующийся таллом представляет собой отдельные клетки, покрытые стенкой, или распадающиеся короткие цепочки клеток (псевдомицелий), которые образуются вследствие нерасхождения материнских и дочерних клеток. Большинство дрожжей размножаются почкованием, при котором новая клетка образуется не вследствие врастания клеточной стенки внутрь клетки, как у мицелиальных грибов, а локальным вздутием оболочки (почка), в которое переходят разделившееся ядро и другие органеллы. Существует также небольшое число «делящихся дрожжей», у которых клетки делятся врастанием оболочки.
БЕЛКИ Структурные белки входят в состав клеточной стенки, мембранных структур, хромосом, из них построены элементы цитоскелета — микротрубочки и микрофиламенты. Ферментные белки обеспечивают все внутриклеточные процессы и взаимодействие с окружающей средой (подробнее см. раздел о питании грибов, с. 272).
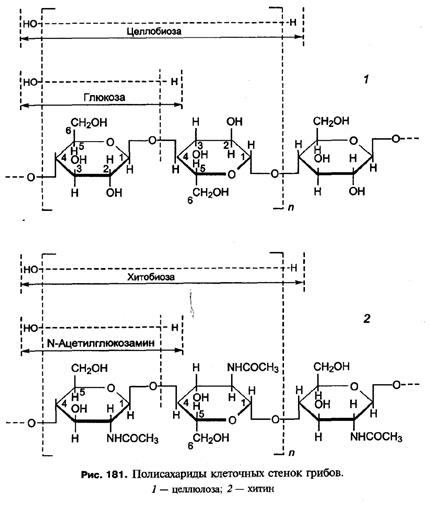
УГЛЕВОДЫ Структурные углеводы составляют основу клеточной стенки грибов. По химическому составу их можно разделить на три группы: полимеры глюкозы, полимеры других моносахаридов и полимеры углеводов, ковалентно связанные с пептидами (гликопротеины). Полимеры глюкозы — это глюканы, хитины и целлюлоза. Глюканы представляют собой линейные или разветвленные цепочки молекул глюкозы, соединенные Полимеры других моносахаров (маннозы, галактозы и др.), называемые у высших растений гемицеллюлозой, встречаются не во всех группах грибов. Особенно много полимеров маннозы (маннанов) в клеточных стенках дрожжей. По-видимому, такой состав стенки лучше, чем глюкановый, обеспечивает почкование. Наконец, в клеточных стенках грибов, как и растений, содержится много полисахаридов, соединенных с белковыми молекулами (пептидоглюканы, маннанопротеины и др.). Они формируют срединный слой многослойной клеточной стенки и играют важную роль как в поддержании структурной целостности клеток, так и в ее обменных процессах с окружающей средой. Запасные углеводы. У грибов не обнаружен крахмал — основной запасной полисахарид высших растений и многих водорослей. Глюкоза у эумицетов запасается в качестве
ЛИПИДЫ Липиды (эфиры глицерина с монокарбоновыми кислотами, имеющими не- разветвленную алифатическую цепь) являются важными запасными продуктами, откладывающимися в клетке в виде капелек жира. для грибов характерно высокое содержание полиненасыщенных (имеющих несколько двойных связей в алифатической цепочке) жирных кислот, таких, как линоленовая с тремя и арахидоновая с четырьмя двойными связями. В виде фосфолипидов (соединений эфирной связью с фосфорной кислотой) липиды входят в состав клеточных мембран (как основной компонент). Большую роль в создании мембранных структур играют также липоиды стерины, придающие мембранам прочность. В отличие от холестерина животных, имеющего 27 атомов углерода в молекуле (С-27), и фитостеринов растений (С-29), основной стерин грибов — эргостерин (С-28). ВТОРИЧНЫЕ МЕТАБОЛИТЫ Пигменты. Грибы лишены фотосинтетических пигментов, но продуцируют большое число соединений, окрашивающих мицелий, пропагативные органы или субстрат. По химической природе большинство пигментов относится к терпеноидам (каротиноиды) или к ароматическим соединениям. Пигменты выполняют разнообразные функции. Так, оранжевые производные каротина индуцируют протекание полового процесса у мукоровых грибов; темно-зеленые и черные фенольные пигменты аспергиллов откладываются только в спороносном аппарате, который, в отличие от субстратного мицелия, образуется в воздушной среде, и в спорах для защиты от ультрафиолетового света; темноокрашенный меланин откладывается в клеточных стенках, что повышает их прочность. Токсины и антибиотики. Многие грибы образуют соединения, токсичные для других организмов. Вещества, токсичные для микроорганизмов, называют антибиотиками, токсичные для растений — фитотоксинами, токсичные для животных и человека — микотоксинами. Некоторые метаболиты грибов, будучи токсичными для разных групп организмов (микроорганизмов и растений, растений и животных), имеют комплексное действие. Антибиотиками обладают многие почвообитающие грибы, которым приходится конкурировать за питательные субстраты с другими микроорганизмами. Их химическая природа и механизм действия многообразны. Так, антибиотики пенициллины и цефалоспорины ингибируют синтез клеточной стенки у бактерий, трихотецины — синтез белка у эукариотных микроорганизмов, гризеофульвин — митоз. Фитотоксины, выделяемые грибами в ткань зараженного растения, вызывают гибель растительных клеток, которые после этого становятся легкой добычей паразита. Токсины ингибируют ферментативные процессы в клетках зараженных растений (например, тентоксин гриба Аlternariа ингибирует фотосинтетическое фосфорилирование); обладают сильным мембранотропным эффектом и влияют на транспорт веществ через мембраны, трансмембранный перенос ионов (фузариевая кислота, фузикокцин и др.). Микотоксины разделяют на две группы: токсины микроскопических грибов микромицетов и токсины грибов макромицетов, имеющих крупные плодовые тела. Первые особенно опасны у грибов, заражающих растительную продукцию, которая используется в пищу. Например, в склероциях спорыньи накапливаются алкалоиды (гетероциклы, содержащие азот), являющиеся нервно-паралитическими ядами. Они не разрушаются при хлебопечении, поэтому хлеб, выпеченный из муки с примесью размолотых склероциев, крайне опасен. Его использование
может вызвать серьезные отравления, часто с летальным исходом. Другой паразит злаков — возбудитель фузариоза колоса гриб Fusarium — выделяет в зерно терпеноидные токсины, также вызывающие серьезные отравления (хлеб, выпеченный из зараженной фузариозом муки, в народе называли «пьяный хлеб», так как он вызывал головокружение, рвоту и другие симптомы, напоминающие сильное алкогольное отравление). Очень опасен токсин желтого аспергилда — афлатоксин. Его продуцент в странах с теплым климатом поражает различную сельскохозяйственную продукцию, использование которой в пищу или на корм скоту может вызвать серьезные поражения печени и других внутренних органов, часто с летальным исходом. Токсины макромицетов опасны для людей, увлекающихся сбором съедобных грибов, но не умеющих отличить съедобные виды от ядовитых. Токсичность некоторых грибковых соединений обусловлена тем, что они подавляют иммунную систему животных или растений, т.е. являются иммунносупрессорами. По химической природе большинство таких соединений являются циклическими пептидами (замкнутые кольца, образоваяные соединением аминокислот друг с другом). В отличие от белков, циклические пептиды, во-первых, низкомолекулярны (образованы из 5—11 аминокислот) и, во-вторых, соединены друг с другом не матричным синтезом на рибосомах, а с помощью специальных ферментов — циклаз. Из подобных соединений наиболее изучен циклоспорин, продуцируемый грибом Tolypocladium. Его используют в хирургической практике — при пересадке органов для подавления иммунных реакций отторжения чужеродного органа. Фитогормоны. Некоторые грибы, являющиеся паразитами растений, выделяют в зараженную ткань низкомолекулярные соединения, регулирующие ростовые процессы у растений (деление и растяжение клеток, открывание и закрывание устьиц и др.). Таковы терпеноиды гиббереллины, образуемые сумчатым грибом Gibberella. Наследственность Геном. Геномом организма называют все его внутриклеточные структуры, способные к саморепликации и несущие ту или иную наследственную информацию. В грибной клетке к таким структурам относятся ядра (ядерный геном), митохондрии (митохондриальный геном), плазмиды. • Ядерный геном у грибов по своему размеру занимает промежуточное положение между геномом бактерий и высших эукариот. Наприм   Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|



 Коровый слой образован очень плотным сплетением мелких гиф с толстыми оболочками, пропитанными темным пигментом меланином, который придает клеткам особую прочность. Внутренний слой представлен рыхлым сплетением крупных гиф с тонкими оболочками и запасом питательных веществ (каплями жира и др.). Он служит для питания проростков склероция. Ложные склероции не имеют внутренней слоистости. Все их клетки одинаковые и содержат питательные вещества и меланин в оболочке. Склероции могут быть образованы только грибными гифами или же в их образовании принимают участие ткани зараженного хозяина, которые мумифицируются. Таковы, например, мумифицированные, черные с поверхности (вследствие отложения меланина) яблоки зараженные возбудителем плодовой гнили Monilia cinerea, или гусеницы, зараженные грибами из рода Cordyceps.
Коровый слой образован очень плотным сплетением мелких гиф с толстыми оболочками, пропитанными темным пигментом меланином, который придает клеткам особую прочность. Внутренний слой представлен рыхлым сплетением крупных гиф с тонкими оболочками и запасом питательных веществ (каплями жира и др.). Он служит для питания проростков склероция. Ложные склероции не имеют внутренней слоистости. Все их клетки одинаковые и содержат питательные вещества и меланин в оболочке. Склероции могут быть образованы только грибными гифами или же в их образовании принимают участие ткани зараженного хозяина, которые мумифицируются. Таковы, например, мумифицированные, черные с поверхности (вследствие отложения меланина) яблоки зараженные возбудителем плодовой гнили Monilia cinerea, или гусеницы, зараженные грибами из рода Cordyceps.

 - или
- или  -связями. Они составляют наружный слой клеточной стенки большинства грибов. В молекулах хитина глюкозные остатки соединены с аминогруппами (аминированы), к которым в свою очередь присоединены остатки уксусной кислоты (ацетилированы). Длинную неразветвленную
-связями. Они составляют наружный слой клеточной стенки большинства грибов. В молекулах хитина глюкозные остатки соединены с аминогруппами (аминированы), к которым в свою очередь присоединены остатки уксусной кислоты (ацетилированы). Длинную неразветвленную  n-го числа таких модифицированных молекул глюкозы (N-ацетил-глюкозо- амина) называют хитином (рис. 181, 2). Его цепочка составляют внутренний слой грибной клеточной стенки и придают ей жесткость, ригидность, заменяя целлюлозу, которая у большинства грибов отсутствует. Степень ацетилирования молекул глюкозы в хитинах из разных организмов может быть различной. Деацетилированный хитин получил название хитозана. Он встречается лишь у некоторых грибов. Наконец, целлюлоза представляет собой пучок линейных молекул
n-го числа таких модифицированных молекул глюкозы (N-ацетил-глюкозо- амина) называют хитином (рис. 181, 2). Его цепочка составляют внутренний слой грибной клеточной стенки и придают ей жесткость, ригидность, заменяя целлюлозу, которая у большинства грибов отсутствует. Степень ацетилирования молекул глюкозы в хитинах из разных организмов может быть различной. Деацетилированный хитин получил название хитозана. Он встречается лишь у некоторых грибов. Наконец, целлюлоза представляет собой пучок линейных молекул