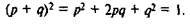|
|
Глава 2 СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИОдной из наиболее уязвимых сторон теории Дарвина было незнание природы наследственности и законов ее изменения. Мы видели, что из всех возражений, адресованных этой теории, самым непреодолимым явилось соображение Дженкина. Оно было столь фундаментальным, что казалось способным дискредитировать всю теорию. Дарвинизм остро нуждался в поддержке генетики, но в течение двух первых десятилетий XX в. ее главные творцы оказались в лагере антидарвинистов, полагая, что генетика сама без всякого отбора в состоянии объяснить эволюцию и видообразование. Не только дарвинизм, но и эволюционная идея в целом были ввергнуты в состояние глубокого кризиса. Однако рано или поздно дарвинизм и генетика в силу логики развития этих наук должны были объединиться. Исторически было предрешено, чтобы плодом их союза стала новая дисциплина — по-пуляционная генетика. Краткая история становления Отправным пунктом популяционной генетики стало правило, или закон Харди—Вайнберга (1908), согласно которому в отсутствие возмущающих факторов (мутаций, миграций, инбридинга, естественного отбора, случайного генетического дрейфа) пропорции генотипов (и, следовательно, частоты генов) в бесконечно большой панмиксической популяции {со свободным скрещиванием) стабилизируются уже в первом поколении, и достигнутое равновесие сохраняется в течение неограниченного числа поколений. Если обозначить через р и q частоты двух аллелей одного гена, то их распределение в такой популяции соответствует коэффициентам разложения бинома Ньютона:
Но такие идеальные популяции — всего лишь абстракция, поскольку в реальной среде постоянно действуют факторы, нарушающие равновесие генотипов. Равновесие популяции — это биологический аналог идеального газа в физике, удобная исходная позиция для определения величины сдвигов равновесия при воздействии различных факторов. Основная заслуга в построении математических моделей действия этих факторов принадлежит англичанам Р. Фишеру и Дж. Б.С. Холдейну и американцу С. Райту. Их трудами в течение немногим более двух десятилетий была создана математическая теория классической популяционной генетики — основы синтетической теории (см. подробнее: Provine, I97I). Уже первые статьи Роналда Фишера (1918, 1922) положили начало использованию в популяционной генетике стохастических методов. Им было открыто явление так называемого сверхдоминирования, при котором если гетерозигота обладает селективным преимуществом, то в популяции между обоими аллелями сохраняется устойчивое равновесие. Целостная концепция эволюционного процесса была разработана Фишером в книге «Генетическая теория естественного отбора» (Fisher, 1930). Она внесла большой вклад в синтез дарвинизма и менделизма и стала для биологов-эволюционистов своего рода Библией. Именно этот труд Фишера более всего способствовал упрочению ортодоксального взгляда, согласно которому характер эволюции определяется почти исключительно естественным отбором. Начало систематическому математическому описанию динамики генных частот под действием естественного отбора положила серия статей Джона Холдейна под обшим названием «Математическая теория естественного и искусственного отбора» (1924-1931). Для определения генных отношений от поколения к поколению им была разработана строгая система нелинейных уравнений. Холдейн установил, что число поколений, необходимое для заданного изменения частоты гена, обратно пропорционально интенсивности отбора, и составил таблицу, отражающую эту количественную закономерность. В последних статьях этой серии было рассмотрено влияние на динамику частоты таких факторов, как неполный инбридинг, ассортативное (избирательное) скрещивание, неполное доминирование в аутосомных и сцепленных с полом генах, сцепление, полиплоидия, изоляция и др. Свои открытия Холдейн подытожил в известной книге «Факторы эволюции» (Haldane, I932; Холдейн, 1936). В 1931 г. в английском журнале «Genetics» вышла большая статья Сьюэлла Райта «Эволюция в менделевских популяциях» (Wright, 1931). В ней было показано, что в основе эволюции лежит процесс смещения популяционного равновесия, в который вовлечены все эволюционные факторы. Среди последних важны не только те, что являются внешними по отношению к популяции, но и связанные с ее собственной организационной структурой. Райт обратил внимание на то, что природные популяции в подавляющем большинстве не являются едиными панмиктическими образованиями, а представляют собой полуизолированные колонии, или субпопуляции, лишь обменивающиеся генетическим материалом. В таких подразделенных и обширных популяциях в результате смешения равновесия процесс эволюции протекает гораздо быстрее. В локальных же субпопуляциях большое эволюционное значение приобретает случайный дрейф генов. Следствием этой закономерности стали выдвинутые много позднее эффект «бутылочного горлышка» и «принцип основателя» (Майр). Работа Райта и указанные труды Фишера и Холдейна фактически завершили в начале 30-х годов XX в. построение классической популяционной генетики, каковой она, по существу, остается до сегодняшнего дня, осуществив тем самым полный синтез дарвинизма и генетики. Стержнем дарвинизма XX в. на Западе стала, таким образом, генетическая теория естественного отбора, представившая последний как статистически-вероятностный механизм. Она предельно упрощала и потому сильно искажала реальную картину динамики популяционных процессов, якобы составляющих начальные этапы эволюции. Так, популяционно-генетическая модель оперировала генами как независимыми единицами, она не учитывала их взаимодействий, игнорировала интегральные эффекты генотипа как целого, возможность изменения генома в ходе индивидуального развития, чрезвычайную сложность связи генотипа с фенотипом и оказалась неспособной объяснить роль в эволюции онтогенетических перестроек. Зато она воодушевляла синтетистов тем, что придавала анализу генетической структуры популяций математическую точность и строгость. Такой путь развития дарвинизма был исторически неизбежным. Синтез генетики и дарвинизма шел не только в рамках теоретико-математических исследований. Он осуществлялся и другими путями. Так, в Советском Союзе, где у истоков синтеза стоял С.С. Четвериков (1926), ведущее значение приобрело изучение природных популяций, данных экспериментальной генетики и действия естественного отбора, во Франции — экспериментальное моделирование с помощью популяционных ящиков, в США — всеми доступными методами. В общей сложности в создании нового синтеза помимо Фишера, Холдейна и Райта приняли участие свыше 30 выдающихся биологов (см. об этом: Галл, Георгиевский, 1973; Галл, Конашев, 1979;. Галл, Георгиевский, Колчинский, 1983; Завадский, Колчин-ский, Ермоленко, 1983). Из их многочисленных трудов первосте- пенное значение приобрели книги О. Добжанского {Dobzhansky, 1937), познакомившего американских эволюционистов со статьей Четверикова, Дж.Г. Симпсона (Simpson, 1944; рус. пер. — 1948), Э. Майра (Мауг, 1942; рус. пер,- 1947) и Дж. Хаксли (Huxley, 1942). От названия последней книги — «Эволюция. Современный синтез» — произошло и название нового синтеза — синтетическая теория эволюции (СТЭ). Впрочем, в западной литературе в большем ходу термин «неодарвинизм». Популяционная генетика и вместе с ней СТЭ сосредоточили свое внимание на исследовании якобы начальных, как считали их создатели, шагов эволюции, завершающихся образованием нового вида и охватываемых понятием микроэволюции. Что касается над-видовой, или макроэволюции, то, по единодушному мнению приверженцев СТЭ, этот уровень эволюции не обладает специфическими факторами и механизмами, отличными от действующих в микроэволюции, и его результаты могут быть выведены из этой последней. Благодаря такому взгляду вся огромная сфера надви-довой эволюции, ради понимания которой эволюционная теория, собственно, и создавалась, фактически выпала из поля зрения син-тетистов. В результате основополагающие понятия СТЭ относятся в основном к микроэволюции» В качестве элементарной эволюционной единицы стали рассматривать популяцию, в качестве элементарной единицы наследственной изменчивости — мутацию, а в качестве элементарного эволюционного явления — изменение генетического состава популяции. К числу элементарных эволюционных факторов в западной литературе были отнесены мутационный процесс, генетический дрейф, изоляция и естественный отбор. В советской науке, следуя за Четвериковым и Тимофеевым-Ресовским, к ним традиционно стали прибавлять «популяционные волны» (см., например: Яблоков, Юсуфов, 1976; Парамонов, 1978). Основные постулаты теории СТЭ восприняла основные положения теории Дарвина и дополнила их новыми. Благодаря ассимиляции данных гораздо большего числа дисциплин и направлений, многие из которых во времена Дарвина просто не существовали, новый синтез предстал в виде обширного собрания эмпирических обобщений, которые ради удобства восприятия желательно было свести к сравнительно небольшому числу тезисов или постулатов. Первым такую попытку предпринял А.А. Любишев (1973), однако ее результат ока- зался нечетким. Вскоре за эту задачу взялся Н.Н. Воронцов, который выделил в СТЭ И основных постулатов (Воронцов, 1978, 1980, 1984), Ниже мы и последуем за Воронцовым, воспроизводя эти постулаты (при этом в ряде случаев отступая от того порядка, в котором они приведены у Воронцова), но сопроводим их краткими комментариями, отвечающими новому видению механизма эволюции. 1. Наименьшей (элементарной) эволюционной единицей яв Как мы имели возможность убедиться, рассмотрение эволюционного процесса исключительно на уровне популяции искажает представления Дарвина и, как мы надеемся показать далее, делает недоказуемым превращение популяции в новый вид, направляя исследование по ложному пути. Популяционная структура вида — очень важная характеристика, обеспечивающая его стабильность и выживание, но прямой связи с эволюцией она не имеет. 2. Материалом для эволюции служат мелкие дискретные наслед Мелкие мутации часто не проявляются на фенотипическом уровне, а те, что проявляются, проходят через «сито» отбора. Кроме мутаций открыты другие типы наследственной изменчивости, с которыми по преимуществу и связана морфологическая эволюция. 3. Основным или даже единственным движущим фактором эво Отбор внутри вида не существует, поскольку, как было сказано в предыдущей главе, внутривидовой борьбы за существование в природе не наблюдается. Без отбора внутри вида нет и начальных шагов эволюции (т.е. микроэволюции), постулируемых в СТЭ. Отбор между разными видами, поскольку он имеет дело с более выраженными фенотипическими различиями, — реальное явление природы, но и эта форма отбора носит относительный характер. 4. Эволюция носит дивергентный характер, т.е. от одного так Поскольку отбор на внутривидовом уровне не работает, не может происходить и дивергенция; коль скоро эти механизмы не работают, исчезает и ограничение, в силу которого новый вид должен был бы возникать только за счет одной популяции. Давно установлено также, что эволюция систематических групп, хотя она и бывает дивергентной, чаще отвечает закону параллельного и конвергентного развития. Многочисленные примеры симгенеза (см. гл. 10) демонстрируют прямо противоположный путь эволюции. 5. Любой реальный, а не сборный таксон имеет монофилети- Этот постулат непосредственно вытекает из предыдущего, и его можно было с ним соединить. В дополнение к соответствующему комментарию следует добавить, что любой вид может совмещать в себе признаки нескольких предковых видов, не говоря уже о высших таксонах, которые, как правило, имеют полифилетиче-ское происхождение. 6. Эволюция носит постепенный (градуалистический) харак Видообразование — одномоментный дискретный акт, совершающийся на протяжении одного или нескольких поколений без всякой подготовки. У видов, размножающихся половым путем, наступление репродуктивной изоляции аналогично беременности: она или происходит, или нет. Представители целого ряда недарвиновских направлений эволюции едины в том, что биологические виды подавляющую часть времени находятся в состоянии стазиса и лишь в самом конце срока своей жизни мутируют, чтобы не прервать нить рода. 7. Обмен генами возможен лишь внутри вида, благодаря че В настоящее время выяснено, что генетическая изоляция видов друг от друга не носит абсолютного характера. Более того, при развитии кризисных ситуаций в экосистемах изолирующие механизмы видов перестают выполнять свою функцию, и в геномы видов поступают чужеродные гены, переносимые различными мобильными генетическими элементами. Такие моменты свободного обмена генами, осуществляющегося неполовым путем, теперь связывают с наиболее революционными событиями в истории биоты. 8. Вид представляет собой сложную полиморфную систему, со Данный постулат не вызывает возражений. Разве что справедливо было бы отметить, что он разделяется далеко не одной СТЭ. 9. Поскольку критерием биологического вида является его По определению этот постулат справедлив. Однако на практике и агамные формы рассматривают как видовые, различая их по морфологическим, физиологическим, генетическим, биохимическим и другим признакам. 10. За рамками концепции биологического вида СТЭ оказа Нам представляется нецелесообразным выделять эту констатацию фактов в отдельный постулат, так как это всего лишь иллюстрация предыдущего постулата. Зато положение о неприменимости понятия вида к ископаемым формам и особенно о том, что эволюция на уровне выше вида идет через микроэволюцию, стоило бы выделить в самостоятельные постулаты. Тезис о сводимости макроэволюции к микроэволюции — один из кардинальных в СТЭ. Он вызвал напряженные и продолжительные дискуссии как в бывшем Советском Союзе, так и за рубежом. Фактически его отрицательное решение уже должно было привести СТЭ к полному банкротству. Этого пока не произошло только в силу сознательного намерения ее приверженцев сохранить данную доктрину любой ценой, не считаясь с фактами. Автор настоящей книги специально посвятил этому фундаментальному вопросу отдельный труд (Назаров, 1991), и мы воспроизведем его наиболее важные разделы в ч. II. II. Исходя из всех упомянутых постулатов, можно сделать заключение, что эволюция непредсказуема, не направлена к некоей конечной цели, следовательно, не имеет финалистического характера. Этот постулат СТЭ также не выдерживает критики. В свете новейших данных законы индивидуального и исторического развития едины, ибо в обоих случаях их объектом выступает биологическая организация, воплощенная в каждой отдельной особи. Онтогенез строго канализован, направлен к конечной цели, заложенной в программе развития, и финалистичен. Каждый таксон, начиная с вида, обладает запрограммированными потенциями к расцвету, процветанию и имеет ограниченный срок жизни (см. гл. 6). Непосредственно наблюдаемые факты целесообразной и коллективной реакции организмов (определенной изменчивости, по Дарвину) на вызов среды дают основание предполагать, что и вся органическая эволюция в целом — целесообразный и направленный процесс. Задача любой теории — его правильно отобразить и, значит, в тех частях, где это логически оправдано, предсказать. Теория, которая не может ни объяснить, ни предсказать, — это плохая теория. Возможно, читая последующие главы, проницательный читатель сможет убедиться, что в объяснении целого ряда закономерностей эволюции и самого феномена жизни лучшие умы человечества исчерпали возможности материалистического подхода и вплотную подошли к признанию верховной власти духовной сферы. После 73-летнего господства в СССР искусственно насаждаемого материализма естествознание робкими шагами постепенно вновь обретает понимание главного источника неслучайности происходящего. И, что самое существенное, можно с удовлетворением констатировать, что это больше не считается антинаучным. Приведенный перечень постулатов, с нашей точки зрения, следовало бы дополнить еще одним очень важным для характеристики СТЭ положением об элементарном эволюционном явлении и поместить его под номером 4. Формулировка этого положения кратка: элементарное эволюционное явление — изменение генетического состава популяции. В соответствии с данным положением все содержание эволюционного процесса стали сводить к отбору и распределению в популяциях мутантных аллелей, часто игнорируя экологические процессы. Такой взгляд на эволюцию получил выражение уже в книге Ф. Добжанского «Генетика и происхождение видов», в которой говорилось, что «изменения генных частот являются наиболее вероятным источником образования... рас», а также видов (Dobzhansky, 1937. Р. 148). Вот и сейчас, через 70 лет, для объяснения возникновения подвидов и видов прибегают к анализу генного состава популяций. О том, решает ли эта процедура проблему, мы поговорим подробнее в последнем разделе главы.   Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|