
|
|
VI. ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ.
В организме одновременно функционирует множество аллельных и неаллельных генов. При этом они могут взаимодействовать друг с другом через биохимические продукты, детерминируемые ими. Основные типы взаимодействия аллельных генов — полное доминирование, неполное доминирование, межаллельная комплементация (кодоминирование) — были разобраны нами в главе «Моногибридное скрещивание». Неаллельные гены также могут взаимодействовать друг с другом, что приводит к изменению их фенотипического проявления. При взаимодействии неаллельных генов расщепление в потомстве по фенотипу будет отличаться от установленных Г.iМенделем. Выделяют следующие типы взаимодействия неаллельных генов: комплементарное, эпистатическое и полимерное. 1. Комплементарность. Комплементарность − это тип взаимодействия неаллельных генов между собой, при котором два неаллельных гена, находясь одновременно в генотипе, дополняют действия друг друга, что приводит к формированию нового признака, отсутствующего у родительских форм. Комплементарность бывает доминантной (когда комплементирующие гены доминантны) и рецессивной (когда формирование нового признака связано с взаимодействием рецессивных аллелей). При доминантной комплементарности присутствие в генотипе доминантных аллелей обоих неаллельных генов обеспечивает развитие нового признака, чего не наблюдается при отсутствии одного из них в доминантном состоянии. В этом случае при скрещивании двух гетерозиготных организмов, имеющих данный (новый) признак, лишь у определенной части потомства (9/16) будет формироваться такой признак, а у остальных (7/16) он не разовьется. Примером доминантной комплементарности может являться наследование окраски цветков душистого горошка. Произведено скрещивание двух сортов душистого горошка с белыми лепестками цветков, у одного из которых были доминантные гены AA и рецессивные bb, а у другого — рецессивные aa и доминантные — BB. Аллель A — отвечает за синтез пропигмента (неактивная форма пигмента), аллель a — не обеспечивает синтез пропигмента, аллель B — отвечает за синтез фермента, который превращает пропигмент в пурпурный пигмент и аллель b — не обеспечивает синтез этого фермента. В потомстве (F1) все растения (генотип АаВb) оказались с пурпурными (красно-фиолетовыми) цветками (новый признак). При скрещивании гибридов первого поколения между собой в F2 получили расщепление по фенотипу: 9/16 растений с пурпурными цветками и 7/16 — с белыми. В целом наследование окраски цветков душистого горошка, иллюстрирующее доминантную комплементарность, может быть представлено в следующей схеме:
Схема 7. Генетическая схема комплементарного взаимодействия неаллельных генов (доминантная комплементарность).
Возможны ситуации, когда каждый доминантный аллель одного гена в отсутствие доминантного аллеля другого неаллельного ему гена (например, А-bb, или ааВ-) обеспечивает развитие «своего» определенного признака, вместе они (А-В-) обусловливает формирование нового варианта признака (доминантная комплементарность), а при наличие в генотипе всех рецессивных аллелей этих неаллельных генов (ааbb) развивается еще один новый вариант признака (рецессивная комплементарность). При этом расщепление в потомстве двух гетерозигот будет составлять 9:3:3:1 или 9:6:1. Таким образом, в этом случае наблюдается и доминантная и рецессивная комплементарность. В качестве примера рассмотрим наследование формы гребня у кур. Доминантный ген А определяет розовидную форму гребня у кур, неаллельный ему доминантный ген В, детерминирует гороховидную форму гребня, при одновременном присутствии в генотипе этих двух доминантных аллелей (А-В-) формируется ореховидная форма гребня, а наличие в генотипе всех рецессивных аллелей данных генов (ааbb) обусловливает развитие листовидной формы гребня. Скрещены две гомозиготные особи с розовидными (ААbb) и гороховидным (ааВВ) формами гребня. Все гибриды (F1) оказались с ореховидной формой гребня (АаВb) (доминантная комплементарность). При скрещивании F1 между собой в потомстве(F2) получили следующее расщепление: 9/16 особей с ореховидным гребнем (А-В-, доминантная комплементарность), 3/16 с розовидной формой гребня (А - bb), 3/16 с гороховидной формой гребня (ааВ-) и 1/16 с листовидной формой гребня (ааbb, рецессивная комплементарность). В целом наследование формы гребня у кур, иллюстрирующее доминантную и рецессивную комплементарность может быть представлено в следующей схеме:
Схема 8. Генетическая схема комплементарного воздействия неаллельных генов (доминантная и рецессивная комплементарность).
Таким образом, при комплементарном взаимодействии генов расщепление по фенотипу в потомстве при скрещивании двух дигетерозиготных организмов будет составлять 9:7 (доминантная комплементарность), 9:3:3:1 или 9:6:1 (доминантная и рецессивная комплементарность). Примерами комплементарного взаимодействия генов у человека могут являться синтез белка интерферона в иммунокомпетентных клетках (определяется взаимодействием двух неаллельных генов), формирование признаков мужского пола (детерминируется несколькими неаллельными генами, определяющими синтез мужского полового гормона — тестостерона и его белка-рецептора). 2. Эпистаз. Эпистаз (от греч. epistasis — остановка, препятствие) - это тип взаимодействия неаллельных генов между собой, при котором ген одной аллельной пары подавляет действие неаллельного ему гена из другой пары. Подавляющий ген называется эпистатическим (супрессором), а подавляемый им ген — гипостатическим. Эпистаз бывает доминантным (когда эпистатический ген представлен доминантным аллелем) и рецессивным, или криптомерия (от греч. kryptos — тайный, скрытый) (когда эпистатический ген представлен рецессивным аллелем). При доминантном эпистазе при скрещивании двух гомозиготных особей, отличающихся по двум парам неаллельных генов, расщепление по фенотипу среди гибридов второго поколения будет выражаться соотношением 12:3:1 или 13:3. При рецессивном эпистазе это соотношение 9:3:4. Примером доминантного эпистаза может служить наследование окраски оперения у кур. У большинства пород кур (плимутрок, виандот) доминантный ген С обусловливает развитие пигментированного оперения, а его аллель с — белого (альбинизм). У породы леггорн существует эпистатический ген I, подавляющий действие доминантного гена С. Его аллель — i не влияет на проявление гена С. Леггорны с генотипом IICC имеют белое оперение в связи с эпистазом гена I над геном С. Виандоты с генотипом iicc имеют белое оперение вследствие отсутствия доминантного аллеля пигментации — С. Проведено скрещивание гомозиготного белого леггорна (IICC) с белым виандотом (iicc). Потомство от этого скрещивания все оказалось белым (IiCc). Далее при скрещивании гибридов первого поколения между собой в F2 наблюдалось следующее расщепление: 13/16 белых кур (I-C-, I-cc, iicc) и 3/16 кур с пигментированным оперением (iiC-). В целом наследование окраски оперения у кур, иллюстрирующее доминантный эпистаз можно представить в схеме 9. Другими примерами доминантного эпистаза могут являться наследование масти у лошадей, окраски плодов тыквы, окраски тела у свиней. Явление рецессивного эпистаза можно проиллюстрировать на примере наследования пигментации шерсти у мышей. Доминантный ген А определяет зональное распределение пигмента по длине волоса у мышей, а его аллель а не влияет на зональное распределение пигмента. Доминантный ген С детерминирует синтез пигмента, а его аллель с определяет отсутствие пигмента (альбинизм) и не дают проявиться гену А (а) т.е. является эпистатическим геном.
Произведено скрещивание гомозиготной черной особи (CCаа) с гомозиготной белой особью (ссАА). Потомство от этого скрещивания казалось серым (СсАа). Далее при скрещивании гибридов первого поколения между собой в F2 отмечено следующее расщепление: 9/16 серых мышей (С-А-), 3/16 черных (С-аа) и 4/16 белых (ссА-, ссаа). В целом наследование пигментации шерсти у мышей, демонстрирующее рецессивный эпистаз можно представить в схеме 10.
Схема 9: Генетическая схема эпистатического взаимодействия неаллельных генов (доминантный эпистаз).
Схема 10. Генетическая схема эпистатического взаимодействия неаллельных генов (рецессивный эпистаз). Примерами рецессивного эпистаза у человека могут являться случаи альбинизма и «бомбейский феномен». «Бомбейский феномен» впервые описан в семье, где у женщины с I группой крови родился ребенок с IV группой крови. Он связан с тем, что в формировании группы крови системы АВО наряду с аллелями гена I (I0, IА, IВ), определяющими синтез антигенов А и В, задействован и ген Н, детерминирующий синтез вещества-предшественника из которого в последующем образуются антигены А и В. У гомозигот по рецессивному аллелю h (hh) даже при наличии в генотипе доминантных аллелей гена I (IА и IВ) антигены А и В не синтезируются, и фенотипически эти люди имеют I группу крови. 3. Полимерия. Полимерия (от греч. polymeria — многосложность) —это тип взаимодействия неаллельных генов между собой, при котором несколько неаллельных генов оказывают влияние на формирование одного признака. Признак не развивается лишь в том случае, когда все полимерные гены находятся в рецессивном состоянии. Одного доминантного гена уже достаточно, чтобы признак развился. Обычно, чем больше доминантных полимерных генов содержит организм, тем сильнее выражен признак. Полимерия лежит в основе наследования количественных признаков. Чем большее число неаллельных генов влияет на количественный признак, тем более плавные переходы этого признака от одного значения к другому. Полимерные гены обозначаются обычно одной и той же буквой, но с разными индексами. Например, генотип включающий три пары неаллельных полимерных генов можно обозначить как — А1а1А2а2А3а3. При полимерии при скрещивании двух гомозиготных особей, отличающихся по двум парам полимерных неаллельных генов, расщепление по генотипу среди гибридов второго поколения будут выражаться соотношением 15:1. Примером полимерии может являться наследование окраски зерен пшеницы. У пшеницы полимерные гены А1 и А2 определяют красную окраску зерен, а их аллели а1 и а2 – отсутствие окраски. Произведено скрещивание гомозиготных организмов, отличающихся по двум парам неаллельных генов (А1А1А2А2 и а1а1а2а2). Все потомство от этого скрещивания имело окрашенные зерна с промежуточной, между белой и красной, интенсивностью окраски (А1а1А2а2). При скрещивании гибридов первого поколения между собой в F2 наблюдалось следующее расщепление: 15/16 с окрашенными зернами (А1-А2-, А1-а2а2, а1а1А2-) и 1/16 с неокрашенными зернами (а1а1а2а2). В целом наследование окраски зерен пшеницы, демонстрирующее полимерию, можно представить в схеме 11.
Схема 11. Генетическая схема полимерного действия неаллельных генов.
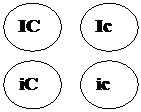
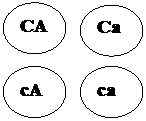
Другими примерами полимерии могут служить наследование формы стручка у пастушьей сумки, яйценоскости у кур, молочности у скота, содержания сахара в корнеплодах, роста и веса животных, длины початка у кукурузы, пигментации кожи, росто-весовых показателей и других количественных признаков у человека. Разберем решение задачи 112. Задача 112. У большинства пород кур (плимутрок, виандот) ген С детерминирует окрашенное оперение, а его аллель с – белое. У породы леггорн имеется эпистатический ген I, подавляющий развитие пигмента даже при наличии гена С. Его аллель – ген i не влияет на проявление гена С. Леггорны с генотипом IIСС имеют белое оперение в связи с эпистазом гена I над С. Виандоты с генотипом iiсс имеют белое оперение вследствие отсутствия доминантного аллеля пигментации – С. Скрещены две белые особи – дигетерозиготный белый леггорн с белым виандотом. Определите расщепление по фенотипу в первом поколении. Запишем условие задачи: Дано:
Выпишем генотипы скрещиваемых особей. Согласно условию задачи это дигетерозиготный белый леггорн (IiСс) и белый виандот (iiсс).
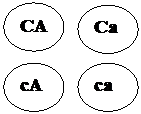
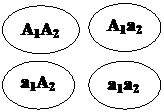
Первый родитель дает четыре типа гамет -,,
гаметы родителей выписываем возможные генотипы потомства и их фенотипы:
  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|

 G
G



 Р2
Р2



 G
G
 F2
Фенотип:
F2
Фенотип:

 G
G



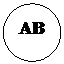



 G
G

 G
G



 G
G
 F2
F2


 F2
F2

 G
G
 G
G
 F2
F2




 и, второй продуцирует один тип гамет -. Комбинируя
и, второй продуцирует один тип гамет -. Комбинируя