|
|
Торможение условных рефлексов
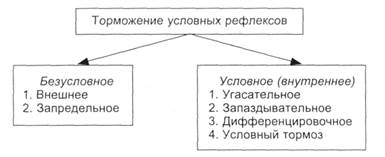
Имеются два вида торможения условных рефлексов, принципиально отличающихся друг от друга: врожденное и приобретенное, каждое из которых имеет собственные варианты (рис. 5.4). Безусловное (врожденное) торможение условных рефлексов подразделяется на внешнее и запредельное торможение. 1. Внешнее торможение – проявляется в ослаблении или прекращении наличного (протекающего в данный момент) условного рефлекса при действии какого-либо постороннего раздражителя. Например, включение звука, света во время текущего условного рефлекса вызывает появление ориентировочно-исследовательской реакции, ослабляющей или прекращающей наличную условно-рефлекторную деятельность. Эту реакцию, возникшую на изменение внешней среды (рефлекс на новизну), И.П. Павлов называл рефлексом «что такое?». Он состоит в настораживании и подготовке организма к действию на случай внезапно возникающей необходимости (нападение, бегство и др.). С повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, поскольку организму не требуется предпринимать каких-то действий. По степени выраженности влияния посторонних раздражителей на условно-рефлекторную деятельность выделяют два варианта торможения: гаснущий тормоз и постоянный тормоз. Гаснущий тормоз – это посторонний сигнал, который с повторением его действия теряет свое тормозящее влияние, поскольку не имеет существенного значения для организма. Обычно на человека действуют ёразличных сигналов, на которые сначала он обращает внимание, а затем перестает их замечать. Постоянный тормоз – это такой дополнительный раздражитель, который с повторением не теряет своего тормозящего действия. Сюда относят раздражения от переполненных внутренних органов (от мочевого пузыря, кишечника и др.), болевые раздражители. Эти раздражители имеют существенное значение для человека и требуют от него принятия решительных мер к их устранению, поэтому условно-рефлекторная деятельность затормаживается.
Рис. 5.4. Виды торможения условных рефлексов
Механизм внешнего торможения. Согласно теории И. П. Павлова, посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, который при средней силе раздражителя оказывает угнетающее влияние на текущую условно-рефлекторную деятельность по механизму доминанты. Внешнее торможение является безусловно-рефлекторным. Поскольку в этих случаях возбуждение клеток ориентировочно-исследовательского рефлекса, возникающего от постороннего раздражителя, находится вне дуги наличного условного рефлекса, это торможение назвали внешним. Более сильный или более важный в биологическом или социальном отношении раздражитель подавляет (ослабляет или устраняет) другую реакцию. Внешнее торможение способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды и дает возможность при необходимости переключиться на другую деятельность в соответствии с ситуацией. 2. Запредельное торможение возникает при длительном нервном возбуждении организма, при действии чрезвычайно сильного условного сигнала или нескольких несильных. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие – «закон силы»: чем сильнее условный сигнал, тем сильнее условно-рефлекторная реакция. Однако закон силы сохраняется до определенной величины, выше которой эффект начинает уменьшаться, несмотря на увеличение силы условного сигнала: при достаточной силе условного сигнала эффект его действия может полностью исчезнуть. Эти факты позволили И. П. Павлову выдвинуть положение о том, что корковые клетки имеют предел работоспособности. Многие исследователи относят запредельное торможение по механизму к пессимальному торможению. Поскольку появление указанного торможения не требует специальной выработки, оно, как и внешние торможения, является безусловно-рефлекторным и выполняет охранительную роль. Условное (приобретенное, внутреннее) торможение условных рефлексов – это активный нервный процесс, который требует своей выработки, как и сам рефлекс. Поэтому его и называют условно-рефлекторным торможением: оно является приобретенным, индивидуальным. Согласно теории И. П. Павлова, оно локализовано в пределах («внутри») нервного центра данного условного рефлекса. Различают следующие виды условного торможения: угасательное, запаздывательное, дифференцировочное и условный тормоз. 1. Угасательное торможение возникает при повторном применении условного сигнала и неподкреплении его. При этом вначале условный рефлекс ослабевает, а затем полностью исчезает, через некоторое время он может восстановиться. Скорость угасания зависит от интенсивности условного сигнала и биологической значимости подкрепления: чем они значительнее, тем труднее совершается угасание условного рефлекса. Этот процесс связан с забыванием полученной ранее информации, если она длительно не повторяется. Если во время проявления условного угасательного рефлекса подействовать посторонним сигналом, возникает ориентировочно-исследовательский рефлекс, который ослабляет угасательное торможение и восстанавливает угасший ранее рефлекс (явление растормаживания). Это показывает, что выработка угасательного торможения связана с активным угасанием условного рефлекса. Угасший условный рефлекс быстро восстанавливается при его подкреплении. 2. Запаздывательное торможение возникает при отставлении подкрепления на 1 – 3 мин относительно начала действия условного сигнала. Постепенно появление условной реакции сдвигается к моменту подкрепления. Более длительное отставление подкрепления в опытах на собаках не удается. Выработка запаздывательного условного рефлекса наиболее трудна. Этому торможению также присуще явление растормаживания. 3. Дифференцировочное торможение вырабатывается при дополнительном включении раздражителя, близкого к условному, и неподкреплении его. Например, если у собаки тон 500 Гц подкреплять пищей, а тон 1000 Гц не подкреплять и их чередовать в течение каждого опыта, то через некоторое время животное начинает различать оба сигнала: на тон 500 Гц будет возникать условный рефлекс в виде движения к кормушке, поедания корма, слюноотделения, а на тон 1000 Гц животное будет отворачиваться от кормушки с пищей, слюноотделения не будет. Чем меньше различия между сигналами, тем труднее идет выработка дифференцировочного торможения. Удается выработать у животных различение частот метронома – 100 и 104 уд./мин, тонов 1000 и 995 Гц, геометрических фигур, а также различение области раздражения различных участков кожи и т.д. Условное дифференцировочное торможение при действии посторонних сигналов средней силы ослабевает и сопровождается явлением растормаживания, т.е. это такой же активный процесс, как и при других видах условного торможения. 4. Условный тормоз возникает при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Так, если выработать условный слюноотделительный рефлекс на свет, затем к условному сигналу «свет» подключить дополнительный раздражитель, например «звонок», и не подкреплять эту комбинацию, то на нее постепенно условный рефлекс угасает. Сигнал «свет» необходимо продолжать подкреплять пищей или вливанием в рот слабого раствора кислоты. После этого присоединение сигнала «звонок» к любому условному рефлексу ослабляет его, т.е. «звонок» стал условным тормозом для любого условного рефлекса. Этот вид торможения также растормаживается, если подключить другой раздражитель. Функциональные изменения при выработке условных рефлексов и условного торможения (изменения возбудимости, ЦНС, ЭЭГ) имеют общие черты, так как стадии их формирования одинаковы. Значение всех видов условного (внутреннего) торможения условных рефлексов заключается в устранении ненужной в данное время деятельности – тонком приспособлении организма к окружающей среде. Механизм условного (внутреннего) торможения условных рефлексов изучен недостаточно (зарегистрирована гиперполяризация клеточных мембран корковых нейронов). При развитии условного торможения в этот процесс широко вовлекаются различные мозговые структуры. Е. Н. Соколов находит много общего в павловском угасательном торможении и в динамике угашения ориентировочного рефлекса. Ученый полагает, что в основе угашения лежит механизм усиления тормозящих влияний на ранее возбуждавшиеся клеточные элементы системы условного рефлекса. Л.Л. Воронин и Р.Г. Кожедуб в модельных ситуациях обнаружили, что при угасательном торможении амплитуда ВПСП постепенно снижалась без заметного перехода в гиперполяризацию клеточной мембраны. Появилась возможность объяснить ослабление возбуждающих реакций уменьшением выброса медиатора без активного торможения самого нейрона. Установлено постепенное угнетение клеточных реакций на угашаемый раздражитель. На клеточном уровне это, по-видимому, осуществляется с помощью механизмов памяти. Полагаем, что торможение условных рефлексов можно объяснить тем, что при неподкреплении условного раздражителя исключаются доминантные взаимоотношения двух очагов возбуждения, остается только возбуждение от условного сигнала. Поэтому проторенный ранее путь между доминантными очагами возбуждения постепенно устраняется, так как исчезает синаптическое облегчение, сформированное ранее. Формы научения
Существует несколько классификаций научения. В частности, все виды научения делят на две группы: неассоциативное (привыкание, сенситизация, импринтинг и подражание) и ассоциативное. Неассоциативное научение реагировать или не реагировать на раздражитель без использования сигнала, без связи (без ассоциации), т.е. без совпадения с каким-то сигналом. Ассоциативное – научение при совпадении индифферентного раздражителя с деятельностью организма. При этом формируется связь (ассоциация) между сигналом и раздражителем (подкреплением). Кроме того, целесообразно все формы научения объединить в четыре основные группы преимущественно по критерию активности животного или человека в ходе научения: 1) пассивное (реактивное) научение; 2) оперантное научение (от лат. operatic – действие); 3) научение с помощью мышления (когнитивное, рассудочное); 4) научение путем инсайта. А. Пассивное (реактивное) научение имеет место во всех случаях, когда организм пассивно, т.е. не прилагая целенаправленных усилий, реагирует на какие-то внешние факторы и когда в нервной системе формируются новые следы памяти. Пассивным научением являются следующие формы. 1. Привыкание – угасание ориентировочной реакции (рефлекса «что такое?» по И. П. Павлову). Если раздражитель многократно повторяется и не имеет особого значения для организма, то организм прекращает на него реагировать, таким образом, развивается привыкание (габитуация). Ориентировочная реакция угасает. Например, появление звука в лаборатории, где находится подопытное животное (собака), первоначально вызывает у него ориентировочную реакцию: поворот головы, ушей в сторону источника звука. После многократного повторения звука, не сопровождающегося другими воздействиями на животное, оно прекращает на него реагировать. У человека и животных новый раздражитель вызывает не только соматические, но и вегетативные реакции: изменение частоты сердечных сокращений, десинхронизацию ЭЭГ, изменение частоты и глубины дыхания. Если оказывается, что раздражитель не имеет значения для организма, то при последующем его повторении угасают и вегетативные, и соматические реакции. Так, человек, живущий на шумной городской улице, не обращает внимания на посторонний шум транспорта ночью и крепко спит. Привыкание – это не только самая простая, но, вероятно, и самая распространенная форма научения у человека и животных. За счет него нам удается игнорировать раздражители, не несущие никакой новизны и не имеющие для нас значения, сосредоточивая внимание на более важных явлениях. Привыкание всегда специфично в отношении стимула: если человек не замечает уличного шума, он проснется либо от неожиданного звука на его фоне, либо от внезапно наступившей тишины. Привыкание – это не утомление, а особый приспособительный нервный процесс. Его не следует путать и с адаптацией анализаторов, т.е. снижением их чувствительности при непрерывной стимуляции. Согласно мнению Е.Н. Соколова, привыкание – это стимул-зависимое обучение, являющееся универсальной формой простейшего отрицательного условного рефлекса, в котором условным сигналом может быть любой раздражитель, а безусловным – биологическая безразличность (подкреплением в этом случае выступает внутреннее состояние, соответствующее отсутствию раздражителя), т.е. привыкание формируется по механизму торможения условных рефлексов (угасательное торможение). На клеточном уровне привыкание формируется с помощью механизмов памяти, как вырабатываются и сами условные рефлексы. 2. Сенситизация – усиление реакции организма на повторяющийся стимул, если он вызывает каждый раз неприятные ощущения. Например, неоднократное капание воды из крана и многократно повторяющееся жужжание назойливой мухи или пронзительный писк комара становятся непереносимыми, неприятными. В данном случае научение носит негативный характер и выражается в соответствующих поведенческих реакциях типа стимул-ответ (после нескольких повторений действия раздражителя). В основе механизма этого вида научения лежит механизм синаптического облегчения. Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей. Кратковременная активация увеличивает амплитуду постсинаптических потенциалов. Облегчение наблюдается и во время раздражения (вначале), в этом случае феномен называют тетанической потенциацией. Степень выраженности облегчения возрастает с увеличением частоты импульсов, оно максимально, когда импульсы поступают с интервалом в несколько миллисекунд. Длительность посттетанической потенциации зависит от свойств синапса и характера раздражения: после одиночных стимулов она выражена слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. По-видимому, главной причиной возникновения синаптического облегчения является накопление Са2+ в пресинаптических окончаниях, поскольку Са2+, который входит в нервное окончание во время ПД, накапливается там, так как ионная помпа не успевает выводить его из нервного окончания. Соответственно увеличивается высвобождение медиатора при возникновении каждого импульса в нервном окончании, возрастает ВПСП. Кроме того, при частом использовании синапсов ускоряется синтез медиатора и ускоряется мобилизация пузырьков медиатора, напротив, при редком использовании синапсов синтез медиаторов уменьшается (важнейшее свойство ЦНС). Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах. Значение синаптического облегчения заключается в том, что оно создает предпосылки улучшения процессов переработки информации на нейронах нервных центров, что крайне важно, например для обучения в ходе выработки условных рефлексов. Повторное возникновение явлений облегчения в нервном центре может вызвать переход центра из обычного состояния в доминантное. 3. Импринтинг – запечатление в памяти новорожденного окружающей действительности. Это – особая форма неассоциативного научения, основанная на врожденной предрасположенности к определенным сочетаниям раздражителей и возникающих ответных реакций в ранний период развития организма. При наблюдениях за животными этологов особенно интересует относительная роль в их поведении врожденного и приобретенного. Работы австрийского этолога Конрада Лоренца помогли понять взаимодействие между этими двумя факторами в некоторых явлениях. В частности, К. Лоренц занимался изучением гусят, вылупившихся в инкубаторе. Первым движущимся объектом, с которым встречались гусята в момент вылупления, была не их биологическая мать, а сам К. Лоренц. Произошло удивительное явление: вместо того чтобы присоединяться к стаду гусей, эти гусята повсюду следовали за К. Лоренцом и вели себя так, как если бы он был их матерью (рис. 5.5). Оказавшись в присутствии своей настоящей матери, они не обращали на нее никакого внимания и возвращались под защиту К. Лоренца. Проявление этой привязанности к человеку стало особенно необычным, когда, достигнув половой зрелости, эти гуси принялись искать брачных партнеров, сходных с человеком, не проявляя ни малейшего интереса к представителям собственного вида.
Рис.5.5. Привязанность к Лоренцу – объекту импринтинга, который гусята увидели после вылупления, заставляет их относиться к нему как к своей настоящей матери [Годфруа Ж., 1992]
К. Лоренц назвал эту глубокую привязанность к первому движущемуся объекту, который увидели гусята после вылупления из яйца, импринтингом (запечатлением). Другие исследователи показали, что в условиях эксперимента импринтинг может быть вызван любым объектом: мячиком для пинг-понга, футбольным мячом, подушкой, картонной коробкой или животным, относящимся к иному виду, при условии, что этот объект движется. Механизм импринтинга важен для выживания. В природных условиях первый движущийся объект, попавший в поле зрения гусят, это обычно их мать; естественно, что импринтинг у них направлен именно на нее и она становится той моделью, которая дает им возможность адекватно проявлять формы поведения, присущие данному виду. Такие явления, хотя они продемонстрированы главным образом у выводковых птиц, у некоторых рыб и ряда млекопитающих, по-видимому, существуют и у птенцовых птиц. У обезьян, т. е. животных, у которых детеныши гораздо дольше зависят от родителей, импринтинг происходит намного позднее и выражен сильнее. У ребенка социальные связи устанавливаются очень рано и носят более глубокий характер. Если индивид первые годы жизни находится в изоляции, то это приводит к отклонениям (иногда значительным) в его поведении. Различают несколько разновидностей (форм) проявления импринтинга. 1. Запечатление образов и объектов: родителей, братьев, сестер, вида пищи и т.д. 2. Усвоение поведенческих актов (дети повторяют действия родителей). Это так называемое имитационное поведение, разновидностью этого импринтинга является половой импринтинг – половое поведение особей внутри своего вида. Но половое поведение у животных может быть направлено и на особь другого вида, с которым воспитывался детеныш. Например, половое поведение кота будет направлено не на кошку, а на собаку, если кот с момента рождения «воспитывался» в семье собаки. 3. Реакция следования – «слепое» (автоматическое) следование новорожденного за родителями. Эта форма импринтинга наиболее демонстративна и хорошо изучена. Считают, что моторным эквивалентом реакции следования животных у ребенка является улыбка, постоянная смена мимики, появление комплекса оживления уже на 2-м месяце жизни. Импринтинг имеет сходство с безусловным и условным рефлексами: а) подобно безусловным рефлексам эти реакции сохраняются, как правило, всю жизнь; б) реакции являются врожденными, но для их проявления нужны определенные условия. Например, реакция следования является врожденной, но в процессе жизни превращается в приобретенную (так, если изолировать детеныша от родителей сразу после рождения на определенный промежуток времени, то реакция следования не формируется, т. е. сама конкретная реакция по наследству не передается, имеется только готовность к ее выполнению у новорожденного). Импринтинг отличается и от условных, и от безусловных рефлексов, а) импринтинг проявляется в определенные критические периоды (чаще сразу или после рождения, причем в короткие сроки, иногда сроки могут быть отдалены); б) запечатление происходит очень быстро, иногда с первого раза, например реакция следования, которую иногда называют условным рефлексом «с одного подкрепления», что, с нашей точки зрения, неверно, так как условный рефлекс надо вырабатывать, а для безусловного рефлекса ни запечатления, ни выработки не требуется. Наблюдение за развитием детей свидетельствует о том, что важную роль в нервно-психическом развитии ребенка имеет импринтинг, влияющий на будущее поведение, предопределяя его на долгие годы. По-видимому, в развитии детей имеются критические периоды, определяющие дальнейшее развитие ребенка, но эти периоды, очевидно, сдвинуты на более поздние сроки, чем у животных. В частности, существует мнение, что возраст от шести недель до шести месяцев является критическим для формирования отношений ребенка с матерью. У новорожденных детей первых месяцев жизни определяющим фактором в возникновении привязанности к матери является чувство комфорта и ощущение безопасности, существенное значение имеют кормление матерью своего ребенка, гигиенический уход, интонация голоса и звуки речи, обязательные при общении с бодрствующим ребенком грудного возраста. Таким образом, импринтинг – это особая, третья форма реагирования (кроме условного и безусловного рефлексов) на окружающую среду. 4. Классические условные рефлексы по И. П. Павлову – положительные и отрицательные (см. разделы 5.2.4 и 5.2.5). Оперантное научение (от лат. operatio – действие) – это научение, в ходе которого организм добивается полезного результата с помощью активного поведения. Имеется два основных подобных вида научения – метод проб и ошибок и инструментальный условный рефлекс. 1. Метод проб и ошибок. Американский ученый Э. Торндайк (1890), видный представитель направления бихевиоризма (от англ. behaviour – поведение) помещал голодных кошек в так называемые проблемные клетки, которые открывались в том случае, если кошка предпринимала какие-то определенные действия: тянула за веревку, приподнимала запорный крючок и т. д. Когда кошка выходила из клетки, она получала пищу. По мере повторения процедуры выхода из клетки (увеличения числа проб и ошибок) скорость выполнения задачи возрастала. Эти исследования были продолжены Скиннером. 2. Инструментальный условный рефлекс – научение действию с помощью вознаграждения (подкрепления). Животное (крыса) по световому сигналу нажимает на рычаг и выключает электрический ток, чтобы избежать раздражения. Крыса также научается реагировать на световой раздражитель: нажимает на рычаг, чтобы получить пищу (рис. 5.6), т.е. она использует какой-то инструмент, отсюда и название этого вида научения. Дети быстро учатся говорить, когда родители одобряют их при правильном произнесении отдельных звуков и слов. В случае неправильного произнесения слова дети не получают подобного подкрепления, в результате чего эти слова постепенно исчезают из употребления в результате неподкрепления. Научение с помощью инструментального условного рефлекса происходит по сигналу, а научение методом проб и ошибок – без сигнала.
Стимулятор и самописец Рис. 5.6. Выработка инструментального условного рефлекса в камере Скиннера. Когда стимулятор автоматически предъявляет раздражитель (здесь – свет), животное может нажать на рычаг и получить пищевое вознаграждение (подкрепление). Раздражители и реакция регистрируются самописцем в виде кривой научения:на абсциссе – день с начала обучения; на ординате – процент правильных реакций на раздражитель
3. Самораздражение структур мозга для получения удовольствия, положительных эмоций или избегание самораздражения (см. раздел 12.3). Научение с помощью мышления (когнитивное научение). Во-первых, имеется в виду такой вариант, когда организм оказывается в ситуации, с которой он ранее не встречался, и находит правильное решение в результате мышления. При условном рефлексе по сигналу тоже имеет место мышление, т. е. оценка результата действия, но это происходит после выполнения действия. Однако в научении с помощью мышления первично мышление, а потом – действие, так как это целенаправленное научение. В эту форму научения следует включить научение с помощью рассудочной и психонервной деятельности, признавая, что между ними различий нет. Во-вторых, в эту форму научения следует включить и научение путем наблюдения. Научение путем наблюдения. Различают два вида подобного научения. 1. Простое подражание. Например, обезьяна в общении с исследователями в экспедиции научилась мыть банан перед едой, не понимая, зачем это делается. 2. Викарное научение, которое также осуществляется посредством наблюдения, но при этом результат действия оценивается. Такое научение свойственно только человеку. Особенно часто используют научение путем наблюдения (подражания) дети, причем в раннем онтогенезе оно является преимущественно просто подражательным. С возрастом викарное научение начинает все больше превалировать над подражательным научением. Экстраполяционные рефлексы, описанные Л. В. Крушинским (1960), состоят в способности животных определять направление дальнейшего перемещения значимого для него раздражителя, например, перемещение пищи, движущейся по прямой линии после исчезновения ее из поля зрения. Способность к экстраполяции хорошо развита у птиц, хищных млекопитающих, дельфинов. Как отмечал Л.В. Крушинский, сложное приспособительное поведение животных строится из трех основных компонентов ВИД: инстинктов, условных рефлексов и рассудочной деятельности, благодаря которой организм сразу вырабатывает программу адаптивного поведения в новых условиях, внезапно сложившихся ситуациях и способен выбрать наиболее адекватную тактику поведения в новой обстановке, прогнозируя развитие событий. Психонервная деятельность. В 1935 г. появилась концепция И.С. Бериташвили, из которой следует, что у высших позвоночных животных и у человека имеется особая форма деятельности мозга – психонервная деятельность, или образная память. Суть психонервной деятельности состоит в том, что у животных возникает определенный образ при однократном восприятии конкретных явлений окружающей действительности (пищи, врага и т.д.), т.е. всех тех объектов внешней среды, которые имеют какую-либо связь с биологически важным состоянием животного (кормлением, самозащитой или размножением). Такой тип поведения включает информационный (образ) и мотивационно-эмоциональный компоненты. Целостные «представления» о среде животным приобретаются при первом же попадании в данную среду, в процессе ее активного ориентировочно-исследовательского освоения. Следует, однако, заметить, что в научении, описанном И.С. Бериташвили и Л. В. Крушинским, различий нет, причем в обоих случаях используется предыдущий опыт. Термины «психонервная деятельность» и «рассудочная деятельность» отражают историю дискуссии о том, думают ли животные. В настоящее время способность животных мыслить не вызывает сомнений. Об этом свидетельствуют, в частности, и экстраполяционные рефлексы, которые описал Л.В. Крушинский. Таким образом, научение с помощью мышления осуществляется, во-первых, путем наблюдения, во-вторых, путем решения задачи в условиях, в которых организм оказывается впервые. Научение путем инсайта (озарения) - внезапное нестандартное правильное решение задачи. Иногда после отдельных проб и ошибок возникает идея результативного действия, которое может совершенствоваться в процессе достижения цели. Этот вид научения является следствием объединения опыта, накопленного в памяти, с той информацией, которой располагает индивид при решении проблемы. Так, находящаяся в клетке обезьяна в эксперименте с недосягаемым для нее бананом сначала пытается достать плод одной палкой в виде трубки, потом другой такой же. Но обе палки были слишком коротки. Тогда она, прекратив свои попытки, стала разглядывать находящиеся возле нее предметы и вдруг быстро проделала логическую последовательность действий: схватила обе палки, вставила их одну в другую, просунула между прутьев клетки и достала банан (рис. 5.7).
Рис. 5.7. Обезьяна путем инсайта находит способ, позволяющий достать далеко расположенный банан с помощью вставленных одна в другую трубок
Следует подчеркнуть, что в конкретных ситуациях для достижения того или иного полезного приспособительного результата индивид чаще всего реализует не один, а несколько видов научения. Частное научение (обучение игре на музыкальном инструменте, обучение работе на компьютере и т.д.) по своей структуре всегда является комплексным, сложным. Большинство форм научения, кроме пассивного и простых (классических, павловских) условных рефлексов, являются сложной формой научения.
5.3. Память* * Раздел 5.3 написан совместно с В. А. Правдивцевым и А.Ф. Беловым.
Общая характеристика
Память – способность организма приобретать, сохранять и воспроизводить в сознании информацию и навыки. Биологическое значение памяти. Накопление, хранение и воспроизведение информации – общие свойства нейронных сетей. Невозможно переоценить биологическое значение этих процессов для адаптации индивидуального поведения к окружающей среде. Без способности к научению и памяти ни отдельная особь, ни вид в целом не могли бы выжить, поскольку оказались бы невозможными как планирование успешных действий, так и преднамеренное избегание ошибок. В последние десятилетия нейробиологи уделяли этим процессам много внимания. Было обнаружено, что запоминается лишь малая часть осознаваемых нами явлений, и они в свою очередь представляют собой ничтожную долю общей сенсорной информации. Однако большинство накопленных сведений со временем, несомненно, забывается. Без таких механизмов отбора и забывания информации мы были бы буквально затоплены ее непрерывным потоком. Классификация. Основу адаптивного индивидуального поведения составляют обучение и память. В нейрологической памяти выделяют генотипическую (врожденную) память, которая обусловливает сохранение инстинктов, импринтинга, и фенотипическую память, мозговые механизмы которой обеспечивают обработку и хранение информации, приобретаемой животным организмом в процессе индивидуального развития. Память различают по формам восприятия информации (логически-смысловая и чувственно-образная, последняя подразделяется на модально-специфические виды – зрительная, слуховая, моторная), по уровням усвоения (воспроизводящая и облегчающая), по длительности хранения информации. Онтогенетическую память составляют условные рефлексы и другие формы приобретенной информации. По длительности хранения информации различные авторы выделяют несколько вариантов памяти, причем везде первым этапом отмечается сенсорная (по мнению некоторых авторов – иконическая) память – доли секунды. Далее начинаются противоречия: а) первичная память – несколько секунд, вторичная – от нескольких минут до нескольких лет, третичная – на протяжении всей жизни; б) кратковременная память – десятки секунд, долговременная – более минуты и на протяжении всей жизни; в) кратковременная (первичная) – десятки секунд, кратковременная (вторичная) – от нескольких минут до нескольких лет, долговременная – на протяжении всей жизни. Общей идеей этих классификаций является длительность хранения информации. Вместе с тем такой подход тоже заключает в себе противоречия. В названных классификациях учитываются не все временные интервалы, их продолжительность не согласуется в разных классификациях. Во-первых, «сенсорную память» необоснованно называют особым видом памяти, так как при этом нет воспроизведения в сознании информации или навыка (воспоминания), а есть продолжение ощущения после выключения раздражителя – как результат инерционности анализаторов, например ощущение прикосновения, которого уже нет. Во-вторых, все противоречия о длительности хранения информации можно исключить, положив в основу классификации не длительность хранения информации, а механизмы: электрофизиологические процессы, биохимические реакции и структурные изменения в нейронах и синапсах ЦНС. При этом идея длительности хранения информации также сохранится – кратковременная, промежуточная и долговременная память. Кроме того, в раннем онтогенезе следует выделить память импринтинга (см. раздел 10.2.6). Механизмы импринтинга связаны с экспрессией в нейронах мозга специфических ранних генов c-fos и c-jun, функцией которых является перестройка работы генетического аппарата нервных клеток под влиянием запечатлеваемого воздействия. По механизму импринтинга у взрослых животных запечатляется действие жизненно значимых подкрепляющих факторов. По мере индивидуального развития животных механизм импринтинга все больше уступает место другим механизмам памяти.
  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|