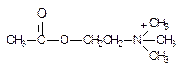|
|
Глава 12. Отравляющие и высокотоксичные вещества нейротоксического действияСтр 1 из 10Следующая ⇒ Глава 12. Отравляющие и высокотоксичные вещества нейротоксического действия
Нейротоксичность - это способность химических веществ, действуя на организм, вызывать нарушение структуры и/или функций нервной системы. Нейротоксичность присуща большинству известных веществ. Она может быть следствием прямого, или опосредованного повреждением других органов, действия токсикантов на нервную систему. Поэтому практически любая острая интоксикация в той или иной степени сопровождается нарушениями функций нервной системы. Тем не менее выделяют группу веществ, условно обозначаемых как нейротоксиканты. К числу нейротоксикантов относят вещества, для которых порог чувствительности нервной системы (отдельных её гистологических и анатомических образований) существенно ниже, чем других органов и систем, и в основе интоксикации которыми лежит поражение именно нервной системы. Токсический процесс, развивающийся в результате действия нейротоксикантов, проявляется нарушением механизмов нервной регуляции жизненно важных органов и систем, а также памяти, мышления, эмоций, поведения и т.д. В основе токсического действия нейротоксикантов может лежать повреждение любого структурного элемента нервной системы путем модификации пластического и энергетического обмена, нарушения генерации, проведения нервного импульса по возбудимым мембранам, передачи сигнала в синапсах. Также условно нейротоксиканты можно разделить на две группы: 1. Вызывающие преимущественно функциональные нарушающие со стороны центрального и периферического отделов нервной системы. 2. Вызывающие органические повреждения нервной системы. Вещества первой группы даже при острых смертельных отравлениях, не вызывают видимой альтерации нервной ткани. Для веществ второй группы, напротив, характерно токсическое действие, сопровождающееся, прежде всего деструктивными изменениями в отделах ЦНС и ПНС. Часть нейротоксикантов вызывают при остром воздействии тяжелые нарушения нервной регуляции функций жизненно важных органов и систем. Для них характерно близкое значение доз, провоцирующих утрату боеспособности (непереносимые дозы) и оказывающих смертельное поражение (смертельные дозы). Поражения этими токсикантами характеризуются большим числом смертельных исходов, а в структуре санитарных потерь преобладают тяжело пораженные, требующие оказания помощи по жизненным показаниям (табл. 38). ОВТВ, обладающие такими свойствами, по большей части относятся к веществам смертельного действия.
Таблица 38. Предполагаемые потери живой силы в районе применения отравляющего вещества VX из выливных авиационных приборов, % (по В.В. Мясникову, 1989)
В основе клиники поражения другими нейротоксикантами лежит преимущественно нарушение функций высшей нервной деятельности. Для основных представителей этой подгруппы характерно более чем тысячекратное различие доз, вызывающих нарушение боеспособности и смерть. Поэтому, как правило, в очагах поражения формируются преимущественно транзиторные токсические реакции и легкие формы отравления, не совместимые с выполнением боевой задачи. Случаи смертельного поражения встречаются редко. Такие вещества относятся к временно выводящим из строя ОВТВ. Из числа представленных в таблице 39 веществ VX, зоман, зарин относятся к первой подгруппе, BZ - ко второй.
Таблица 39. Токсичность некоторых отравляющих веществ (по В.В. Мясникову, 1989)
Большинство промышленных токсикантов, пестицидов, лекарственных средств (применение которых возможно в качестве диверсионных агентов), занимают промежуточное положение между смертельно действующими отравляющими веществами и временно выводящими из строя. Различие значений их смертельных и выводящих из строя доз больше, чем у представителей первой подгруппы, и меньше, чем у представителей второй.
Вещества, вызывающие преимущественно Рисунок 42. Места действия токсикантов на синапс Схема этапов оборота нейромедиатора, воздействуя на которые токсикант может инициировать токсический процесс: 1 - синтез; 2 - хранение; 3 - метаболизм; 4 - выделение; 5 - обратный захват; 6 - разрушение; 7 - рецептор; 8 - механизм проведения сигнала.
Среди отравляющих и высоко токсичных веществ, относящихся к числу нейротоксикантов, вызывающих преимущественно функциональные нарушения, можно выделить две основные подгруппы: 1. Нервно-паралитического действия; 2. Писходислептического действия.
12.1.1. Отравляющие и высокотоксичные вещества нервно-паралитического действия
До недавнего времени к числу веществ нервно-паралитического действия относили исключительно боевые отравляющие вещества из класса фосфорорганических соединений (ФОС) - фосфорорганические отравляющие вещества (ФОВ). В настоящее время запасы ФОВ, хранящиеся в арсеналах, подлежат уничтожению, в связи с чем снижается вероятность их применения при ведении военных действий. Однако появились новые токсиканты, поражение которыми не исключено в ходе военных конфликтов, при аварийных ситуаций на объектах синтеза, хранения высокотоксичных веществ и средствах их транспортировки, при совершении террористических актов. К числу ОВТВ нервно-паралитического действия можно отнести: 1. Фосфорорганические соединения (зарин, зоман, VX, фосфакол, армин, карбафос, дихлофос и др.); 2. Производные карбаминовой кислоты (пропуксор, альдикарб, диоксакарб и др.); 3. Бициклофосфаты (бутилбициклофосфат, изопропилбициклофосфат и др.); 4. Производные гидразина (гидразин, диметилгидразин и т.д.) 5. Сложные гетероциклические соединения (тетродотоксин, сакситоксин, норборнан и др.); 6. Белковые токсины (ботулотоксин, тетанотоксин). Структурные формулы отдельных представителей групп даны на рисунке 43.
Рисунок 43. Строение некоторых представителей ОВТВ нервно-паралитического действия
Современные нервно-паралитические ОВТВ различаются не только строением, но и особенностями токсического действия (таблицы 3). Вещества одной группы при тяжелых интоксикациях вызывают развитие судорожного синдрома, комы и гибели пострадавшего, как правило, от остановки дыхания и сердечной деятельности на фоне истощения энергетических ресурсов организма. Другие – первично вызывают паралич произвольной мускулатуры, в том числе и дыхательной, и гибели от асфиксии. Интоксикация некоторыми веществами развивается достаточно быстро, и у медицинской службы практически нет резерва времени для спасения жизни пострадавших. В этих случаях с целью медицинской защиты личного состава должны использоваться антидоты само- и взаимопомощи. Интоксикация другими представителями группы развивается медленно. Основной задачей врача в этом случае является своевременное выявление пораженных и скорейшая эвакуация на этапы оказания квалифицированной помощи. В связи с этим нервно-паралитические ОВТВ можно классифицировать следующим образом (табл. 40):
Таблица 40. Судорожного действия Рисунок 44. Схема функционирования холинэргического синапса ХР - холинорецептор; АХЭ – ацетилхолинэстераза; 1 - 3 - способы воздействия токсикантов на синапс: 1 - ботулотоксин; 2 - ингибиторы АХЭ; 3 - холинэргические и антихолинэргические средства
Установлено, что нейромедитор и в ЦНС и в ПНС взаимодействует с двумя видами рецепторов: мускариновыми и никотиновыми, отличающимися между собой способностью отвечать на воздействие ряда агонистов (М-холинорецепторы избирательно возбуждаются мускарином; Н-холинорецепторы - никотином), механизмом рецепции сигнала и его передачи на эффекторную клетку (нейрон, миоцит, железистую клетку). Н-холинорецепторы непосредственно связаны с ионными каналами постсинаптической мембраны для Na+, K+, Ca2+. М-холинорецепторы представляют собой структуры, которые также влияют на проницаемость ионных каналов для Na+ и K+. Однако это влияние опосредовано изменением активности сопряженных с рецептором ферментов, регулирующих содержание в иннервируемой клетке вторичных мессенджеров восприятия сигнала (цАМФ, цГМФ, фосфоинозитола и т.д.). И в том и в другом случае действие ацетилхолина на рецепторы приводит к усилению проницаемости постсинаптических мембран для ионов натрия, что сопровождается деполяризацией мембраны и ее возбуждением. Различается скорость генерации постсинаптического сигнала: в Н-холинэргических синапсах изменение конформации ионных каналов развивается практически мгновенно, а в М-холинэргических - постепенно. Установлено, что в синапсах на пресинаптической мембране локализуются холинорецепторы, выполняющие функцию регуляторов проведения импульса. Прерывание нервного импульса осуществляется путем гидролиза ацетилхолина ферментом ацетилхолинэстеразой. Механизм взаимодействия ацетилхолина с холинэстеразой изучен достаточно подробно. Известно, что в состав молекулы ацетилхолина входят реакционноспособные атомы, которые образуют две группы: катионную (четвертичный атом азота с тремя метильными радикалами) и сложноэфирную (эфирный кислород и карбонильные углерод и кислород) (рис. 45).
Рисунок 45. Структура ацетилхолина
Соответственно реакционным группам ацетилхолина, в активных центрах холинэстеразы также выделяются два участка: анионный и эстеразный. Предполагается, что анионный участок образован карбоксил-анионом двухосновной аминокислоты. На расстоянии 0,4-0,5 нм от анионного центра расположен эстеразный участок, в котором функциональную роль играет гидроксильная группа аминокислоты серина. Взаимодействие ацетилхолина с холинэстеразой начинается с ориентации катионной “головки” ацетилхолина на анионном участке фермента. Это приводит к фиксации молекулы ацетилхолина на определенном расстоянии от эстеразного участка, на котором и проходит ферментативный гидролиз медиатора (разрушение эфирной связи между холином и ацетатом). Для его осуществления гидроксильная группа серина проводит нуклеофильную атаку на соответствующий электрофильный атом субстрата. В результате реакции образуются ацетилированный по серину фермент и холин, сорбированный на анионном участке. В последующем холин отщепляется от анионного участка фермента, а затем происходит и деацетилирование холинэстеразы - восстанавливается ее первоначальная структура. В ЦНС плотность М-холинэргических синапсов между нейронами существенно выше, чем Н-холинэргических. Наивысшая - в хвостатом, прилежащем, чичевичном ядрах, гиппокампе, гипоталамусе, коре головного мозга, мозжечке млекопитающих и человека. На периферии М-холинэргические синапсы связывают окончания постганглионарных нервных волокон парасимпатической системы с иннервируемыми ими гладкомышечными волокнами (кишечник, бронхи, миокард) и клетками экзокринных желез (слюнных, желудочно-кишечного тракта, бронхиальных, потовых). Периферические Н-холинэргические синапсы выявляются главным образом в области окончаний нервных волокон мотонейронов, иннервирующих поперечно-полосатую мускулатуру, волокон, иннервирующих ганглионарные нейроны парасимпатического и симпатического отделов вегетативной нервной системы, а также в каротидном клубочке, хромаффонинных клетках мозгового вещества надпочечников. Как указывалось ранее, перевозбуждение центральных холинэргических механизмов может приводить к развитию судорожного синдрома. Наибольшей активностью в этом плане обладают ингибиторы АХЭ, способные проникать через гематоэнцефалический барьер (ФОС, карбаматы). Перевозбуждение периферических холинореактивных структур этими токсикантами сопровождается существенным, порой не совместимым с жизнью, нарушением функций органов и систем.
Ингибиторы холинэстеразы
Рисунок 46. Структура некоторых фосфорорганических соединений
Биологическая активность ФОС, в том числе и токсичность, зависит от их строения (табл. 43).
Таблица 43. Токсичность (ЛД50) некоторых ФОС для белых мышей
Все ФОС обладают высокой реакционной способностью. Особое значение придают реакциям фосфорилирования, гидролиза и окисления, поскольку именно эти реакции определяют стойкость токсикантов в окружающей среде, имеют отношение к метаболизму и механизму токсического действия ядов в организме, на них основаны некоторые принципы дегазации, обнаружения, антидотной профилактики и терапии интоксикаций. ФОС легко отдают электроны, активно вступают в реакции с электрофильными группами других соединений и за счет этого фосфорилируют многие вещества (аминокислоты, полифенолы, гидроксиламин, гидроксамовые кислоты и др.). В качестве примера приводим реакцию фосфорилирования зарином гидроксиламина:
Все ФОС при взаимодействии с водой подвергаются гидролизу с образованием нетоксичных продуктов. Скорость гидролиза ФОС, растворенных в воде, различна (например, зарин гидролизуется быстрее, чем зоман, а зоман – быстрее, чем V-газы). В общей форме реакция гидролиза может быть представлена следующим образом:
Реакция гидролиза ФОС с разрывом ангидридной связи происходит и в организме, как спонтанно, так и при участии энзимов. В результате реакции окисления, ФОС также разрушаются, однако в ряде случаев (при окислении фосфотионатов до фосфатов) некоторые вещества даже повышают свою активность. Это иллюстрируется примером
Токсичность параоксона для млекопитающих и человека выше, чем паратиона. Важнейшие свойства фосфорорганических отравляющих веществ представлены в таблицах 44-46.
Таблица 44. Основные свойства зарина
Таблица 45. Основные свойства зомана
Таблица 46. Патогенез интоксикации Пусковым механизмом практически всех симптомов, развивающихся при интоксикации ФОС, является перевозбуждение никотиновых и мускариновых холинергических синапсов, локализованных в центральной нервной системе и на периферии (табл. 47). При тяжелых поражениях в патологический процесс, по мере развития интоксикации, вовлекаются и нехолинэргические механизмы. К числу таковых относятся: нарушение функционального состояния глутаматэргической, катехоламинэргических, ГАМК-эргической нейромедиаторных систем мозга, увеличение содержания в крови биологически активных веществ (гормонов, продуктов перекисного окисления липидов, лейкотриенов, простогландинов, фактора агрегации тромбоциотв и т.д.), прогрессирующая гипоксия, изменение кислотно-основного состояния и электролитного баланса и т.д.
Таблица 47. Рисунок 47. Схема взаимодействия ацетилхолина, зарина и фосфорилтиохолина с активным центром ацетилхолинэстеразы
Чем выше структурное сходство ФОС с ацетилхолином, тем, как правило, выше его антихолинэстеразная активность и токсичность. Принципиальное различие во взаимодействии ацетилхолина и ФОС с АХЭ состоит в том, что реакция декарбоксилирования активного центра после гидролиза АХ проходит практически мгновенно и энзим снова восстанавливает способность взаимодействовать с субстратом, а дефосфорилирование – протекает медленно. Причем с течением времени изначально обратимая связь ФОС-АХЭ, которая может разрушаться спонтанно (“спонтанная реактивация”) или с помощью некоторых веществ, вводимых отравленному (реактиваторы АХЭ), становится необратимой, неспособной к разрушению. Таким образом, взаимодействие ФОС и АХЭ проходит в две фазы и может быть представлено следующим образом:
Процесс превращения образовавшейся в первой фазе обратимо фосфорилированной холинэстеразы в необратимо связанную форму называется “старение” фосфорилхолинэстеразы. Как скорость “спонтанной реактивации” АХЭ (и самопроизволное восстановление ее активности), так и “старения”, зависит от структуры ФОС, а именно от строения алкильных радикалов при атоме фосфора. Чем “тяжелее” радикалы, тем ниже скорость “спонтанной реактивации” и выше скорость “старения”. Поэтому АХЭ, ингибированная VX (R -OC2H5), стареет чрезвычайно медленно, зарином (R -OCН(СH3)2) - в течение нескольких часов, зоманом (R -OCНСН3С(СH3)3) - в считанные минуты. В основе “старения” лежит процесс отщепления от атома фосфора, связанного с активным центром энзима, алкильных радикалов. При этом одновременно изменяется конформация белковой части энзима (с этим, вероятно, связано то обстоятельство, что фосфорилированные одним и тем же веществом холинэстеразы, выделенные из тканей разных млекопитающих, “стареют” с разной скоростью). В настоящее время обнаружены соединения (гидроксиламин, гидроксамовые кислоты, оксимы), способные, взаимодействуя с остатком ФОС, связанного с АХЭ, отрывать его от молекулы энзима (если не произошло его “старение”) и, тем самым, восстанавливать ферментативную активность. Такие вещества, получившие название реактиваторов холинэстеразы, при своевременном введении отравленному существенно ослабляют выраженность токсического процесса, что подтверждает справедливость антихолинэстеразной теории действия ФОС. Даже ингибированная Vх ацетилхолинэстераза, “стареющая” с минимальной скоростью, а “спонтанно реактивирующаяся” относительно быстро, дефосфорилируется в течение нескольких суток. Вот почему ФОС называют необратимыми ингибиторами холинэстеразы. In vitro способность ФОС угнетать АХЭ уменьшается при увеличении концентрации в инкубационной среде естественного субстрата энзима - ацетилхолина. По этой причине ФОС называют также конкурентными ингибиторами АХЭ. За активный центр энзима ФОС конкурируют не только с ацетилхолином, но и с ингибиторами энзима из других классов соединений, в частности с карбаматами. Последние вызывают обратимое карбамилирование активного центра АХЭ и потому называются обратимыми ингибиторами АХЭ. Установленная в опытах in vitro и in vivo способность обратимых ингибиторов холинэстеразы (прозерина, галантамина и т.д.) защищать холинэстеразу от угнетения ФОС, предупреждать действие этих ядов на органы и системы и, тем самым, препятствовать развитию интоксикации используется на практике при разработке профилактических антидотов ФОС (см. ниже). Холинэстеразная активность выявляется не только в синаптических структурах, но и в крови млекопитающих и человека. Причем в мембране эритроцитов содержится ацетилхолинэстераза, по сути, идентичная энзиму нервной ткани, а в плазме крови - бутирилхолинэстераза, отличающаяся от АХЭ более высоким сродством к эфирам холина и жирных кислот с большей молекулярной массой, чем ацетат (например, бутирилхолину - эфиру холина и масляной кислоты). ФОС, при поступлении в организм, угнетают оба типа холинэстераз крови. Степень инактивации энзимов пропорциональна степени угнетения активности синаптической ацетиохолинэстеразы. Это явление используется для диагностики интоксикации ФОС, а также для верификации степени тяжести поражения. При отсутствии иных причин, снижение активности холинэстеразы крови более чем на 50% свидетельствует об интоксикации антихолинэстеразными ядами. При необходимости контролировать состояние синаптического энзима, для исследования можно изучать активность АХЭ в эритроцитах, выделенных методом центрифугирования. Способность ФОС угнетать холинэстеразу используют также для индикации ФОС в воде, продовольствии и т.д. (биохимический метод индикации). Действие на холинорецепторы. Из возможных неантихолинэстеразных механизмов наиболее важным является действие ФОС на холинорецепторы. Поскольку и холинорецепторы, и холинэстераза адаптированы к одному и тому же нейромедиатору, ингибиторы холинэстеразы могут проявить активность и по отношению к холинорецепторам. По-видимому, блокада проведения нервно-мышечного сигнала, развивающаяся при смертельной интоксикации ФОС, связана не только со стойким деполяризующим действием избыточного количества ацетилхолина, но и с прямым действием ФОС на нервно-мышечные синапсы (по типу действия деполяризующих миорелаксантов). Так, в эксперименте на изолированном нервно-мышечном препарате млекопитающего, при внесении в инкубационную среду достаточной дозы ФОС, наблюдается полное прекращение передачи нервного импульса с нервного волокна на мышцу. Однако через некоторое время на фоне практически “тотального” угнетения активности холинэстеразы отмечается восстановление нервно-мышечной проводимости в синапсах. Повторно блок можно вызвать, вновь добавив ФОС в инкубационную среду. Сенсибилизирующее действие на холинорецептор зарина, ДФФ и других ФОС, проявляется, в частности, существенным повышении чувствительности отравленных экспериментальных животных к холиномиметикам, негидролизуемым ацетилхолинэстеразой (никотину, ареколину и т.д.). Установлено, что сенсибилизация к М-холиномиметикам (ареколину) сохраняется значительно дольше, чем к Н-холиномиметикам (никотину). Причины различия, вероятно, обусловлены особенностями проведения нервных импульсов в М- и Н-холинэргических синапсах (см. выше). Восстановление нормального проведения нервного импульса у лиц, перенесших интоксикацию ФОС, осуществляется за счет медленно протекающих процессов дэфосфорилирования АХЭ (“спонтанная реактивация”), синтеза АХЭ в перикарионе нервных клеток de novo и транспорта ее в нервные окончания, снижения содержания ацетилхолина в синаптической щели, десенситизации холинорецепторов (понижение чувствительности к ацетилхолину). Нехолинэргические механизмы токсического действия. Помимо действия на холинореактивные структуры, ФОС, в высоких дозах, обладают прямым повреждающим действием на клетки различных органов и тканей (нервной системы, печени, почек, системы крови и т.д.), в основе которого лежат общие механизмы цитотоксичности: нарушение энергетического обмена клетки; нарушение гомеостаза внутриклеточного кальция; активация свободнорадикальных процессов в клетке; повреждение клеточных мембран. Чем менее токсично ФОС, тем значимее роль указанных механизмов в развитии проявлений тяжелого поражения данным токсикантом. Существуют ФОС полностью лишенные антихолинэстеразной активности, токсичность которых обусловлена исключительно их цитотоксическим действием (три-о-крезилфосфат). Клиника отравления такими веществами полностью отличается от описанной выше. Медицинские средства защиты К числу средств медицинской защиты относятся: 1. Средства предотвращающие поступления веществ в организм через кожу - индивидуальные противохимические пакеты (ИПП - см. раздел “Специальная обработка”); 2. Медикаментозные средства медицинской защиты. Медикаментозные средства представлены препаратами, назначаемыми с целью профилактики поражения, само- и взаимопомощи, раннего (догоспитального) лечения. Сюда относятся этиотропные, патогенетические и симптоматические средства. Этиотропные средства защиты (антидоты). Разработка этиотропных средств защиты - специфических противоядий (антидотов) ФОВ, долго считавшихся основным видом химического оружия, началась сразу после второй мировой войны и продолжается до настоящего времени. Основные направления исследований в области разработки медицинских средств защиты представлены в таблице 48.
Таблица 48. Рисунок 48. Структура некоторых реактиваторов холинэстеразы
Биохимический механизм процесса реактивации холинэстеразы (Е) схематично представлен на рисунке 49.
Рисунок 49. Дефосфорилирование хлинэстеразы с помощью оксима
В результате нуклеофильной атаки оксимной группы на фосфорилированный энзим разрывается ковалентная связь между атомом фосфора ФОС с кислородом серина активного центра холинэстеразы и образуется новое соединение – фосфорилированный оксим и свободный энзим. Реактивация ингибированных холинэстераз как in vitro, так и in vivo характеризуется двумя показателями: скоростью, то есть количеством восстановленного энзима в единицу времени, и уровнем реактивации – максимальным количеством дефосфорилированного фермента, которое можно получить, обрабатывая фосфорилированную холинэстеразу реактиватором. Эти характеристики зависят от структуры ФОС, строения реактиватора и времени, прошедшего с момента взаимодействия ФОС с активным центром холинэстеразы. Например, скорость реактивации холинэстеразы эритроцитов человека, ингибированной зарином, под влиянием 2 ПАМ в 10 раз выше, чем холинэстеразы, угнетенной ДФФ. Феномен может быть следствием влияния стерических (пространственно-структурных) факторов. Так, при реактивации холинэстеразы, ингибированной веществами с сильно разветвленными алкильными радикалами (ДФФ), затрудняется образование связи между атомом фосфора и кислородом оксимной группы, без чего невозможна реактивация. Из известных реактиваторов наивысшей активностью обладают биспиридиниевые соединения (содержат два пиридиновых кольца в молекуле): ТМБ-4, токсогонин и др. Слабой реактивирующей и антидотной активностью, не смотря на способность проникать через ГЭБ, обладают реактиваторы, не содержащие в молекуле пиридиновых радикалов с четвертичным азотом: диацетиламиноксим (ДАМ-оксим), изонитрозин и др. Как указывалось ранее, холинэстеразы, ингибированные ФОС, с течением времени приобретают устойчивость к воздействию реактиваторов. Это явление получило название “старения” фосфорилхолинэстераз. Поэтому на реактивационный эффект при действии оксимов можно рассчитывать только при условии их применения в первую (обратимую) фазу торможения, продолжительность которой, в зависимости от строения действующих ФОС, будет значительно различаться (минуты – часы). Наиболее устойчивой к реактиваторам является быстро “стареющая” холинэстераза, ингибиорованная зоманом. Помимо свойства дефосфорилировать холинэстеразу и восстанавливать ее активность, реактиваторы способны деблокировать (десенсибилизировать) холинорецепторы и восстанавливать их функцию, разрушать ФОС при непосредственном взаимодействии с ядами. Деблокирующее действие реактиваторов достаточно подробно изучено на мионевральных синапсах. Так, показано, что с помощью 2-ПАМ, ТМБ-4 удается устранять блок проведения на нервно-мышечном препарате диафрагмы крысы, вызванный зоманом или метилфторфосфорилхолином. Этот факт нельзя объяснить реактивацией холиэстеразы, так как яды образуют с ферментом практически нереактивируемый комплекс. Десенсибилизирующее действие оксимов, проявляется способностью снижать возникающую при действии ФОС повышенную чувствительность Н-холинорецепторов (центральной нервной системы, вегетативных ганглиев) к холиномиметикам, не гидролизуемым холинэстеразой. Полагают, что и деблокирующее и десенсибилизирующее действие оксимов на Н-холинорецепторы осуществляется за счет расщепления комплекса ФОС – холинорецептор. Некоторые оксимы (2-ПАМ, ТМБ-4) обладают Н-холинолитической активностью (курареподобным действием), что также играет роль в их способности устранять нервно-мышечный блок. Один из механизмов защитного действия реактиваторов связан с прямой нейтрализацией (разрушением) яда, циркулирующего в крови. Суть явления состоит в том, что оксимы образуют связь с атомом фосфора ингибитора, замещая при этом подвижную группу в молекуле ФОС. При этом образуется комплекс яда с оксимом, который, распадаясь, дает уже неактивное соединение. Однако в некоторых случаях скорость распада продукта взаимодействия ФОС–оксим мала. При этом в организме накапливается вещество, обладающее, порой, мощными ингибиторными свойствами. Поэтому в практике оказания помощи отравленным следует избегать введения неоправданно большого количества оксимов. В последнее время появились сообщения о способности реактиваторов холинэстеразы восстанавливать активность карбоксилэстеразы крови и тканей, что важно, поскольку при отравлении ФОС этот фермент играют существенную роль в детоксикации ядов. Под влиянием реактиваторов холинэстеразы ускоряется восстановление сознания у пораженных, ослабляется выраженность бронхоспазма, спазма кишечника, ослабляются или прекращаются фибриллярные мышечные подергивания, предупреждается развитие мышечной слабости, нормализуется функция нервно-мышечных синапсов и восстанавливается деятельность дыхательной мускулатуры, что в свою очередь приводит к ослаблению гипоксии. Изложенные представления о механизмах антидотного действия оксимов определяют тактику их применения. Необходимостью раннего введения обусловлено включение реактиватора холинэстеразы в состав антидота само- и взаимопомощи АЛ-85 (см. ниже). В качестве лечебных антидотов реактиваторы применяют при отравлениях средней степени тяжести и тяжелых поражениях ФОС. В опытах на животных показано, что минимальной действующей концентрацией пралидоксима (2-ПАМ) в крови является 4 мг/мл. Внутривенное введение препарата со скоростью 500 мг/час обеспечивает поддержание концентрации в плазме крови 15 мг/мл. Для достижения действующей концентрации препаратов в крови и тканях человека и поддержания ее на этом уровне в течение необходимого времени (около 1,5 часов) рекомендуют повторно, с интервалом 20 минут вводить внутривенно (или внутримышечно) 500 - 2000 мг пралидоксима (2-ПАМ) или 150 - 250 мг дипироксима (ТМБ-4). При необходимости инъекции можно повторить спустя 4 - 6 часов после первого введения. Возможно внутривенное капельное введение препаратов в указанных дозах со скоростью: 2-ПАМ - 100 мг/мин, ТМБ-4 - 25 мг/мин. Более высокие дозы оксимов или более частое и длительное их применение не только не оправд   Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|