|
|
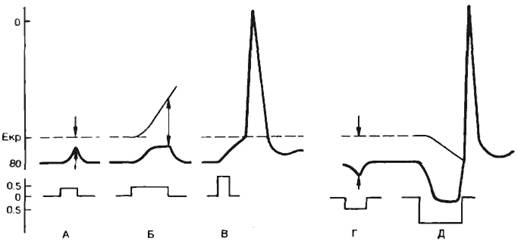
Действие электрического тока на возбудимые тканиЭлектрический ток широко используется в экспериментальной физиологии при изучении характеристик возбудимых тканей, в клинической практике для диагностики и лечебного воздействия, поэтому необходимо рассмотреть механизмы воздействия электрического тока на возбудимые ткани. Реакция возбудимой ткани зависит от формы тока (постоянный, переменный или импульсный), продолжительности действия тока, крутизны нарастания (изменения) амплитуды тока. Эффект воздействия определяется не только абсолютным значением тока, но и плотностью тока под стимулирующим электродом. Плотность тока определяется отношением величины тока, протекающего по цепи, к величине площади электрода, поэтому при монополярном раздражении площадь активного электрода всегда меньше пассивного. Постоянный ток. При кратковременном пропускании подпорогового постоянного электрического тока изменяется возбудимость ткани под стимулирующими электродами. Микроэлектродные исследования показали, что под катодом происходит деполяризация клеточной мембраны, под анодом—гиперполяризация (рис. 2.14, А). В первом случае будет уменьшаться разность между критическим потенциалом и мембранным потенциалом, т. е. возбудимость ткани под катодом увеличивается. Под анодом происходят противоположные явления (рис. 2.14, Г), т. е. возбудимость уменьшается. Если мембрана отвечает пассивным сдвигом потенциала, то говорят об электротонических сдвигах, или электротоне. При кратковременных электротонических сдвигах значение критического потенциала не изменяется. Поскольку практически у всех возбудимых клеток длина клетки превышает ее диаметр, электротонические потенциалы распределяются неравномерно. В точке локализации стимулирующего электрода сдвиг потенциала происходит очень быстро и временные параметры определяются величиной емкости мембраны. В удаленных участках мембраны ток проходит не только через мембрану, но и преодолевает продольное сопротивление внутренней среды. Электротонический потенциал падает экспоненциально с увеличением длины, а расстояние, на котором он падает в 1/е раз (до 37%), называют константой длины (λ). При сравнительно большой продолжительности действия подпорогового тока изменяется не только мембранный потенциал, но и значение критического потенциала. При этом под катодом происходит смещение уровня критического потенциала вверх (рис. 2.14, Б), что свидетельствует об инактивации натриевых каналов. Таким образом, возбудимость под катодом уменьшается при длительном воздействии подпорогового тока. Это явление уменьшения возбудимости при длительном действии подпорогового раздражителя называетсяаккомодацией. При этом в исследуемых клетках возникают аномально низкоамплитудные потенциалы действия.
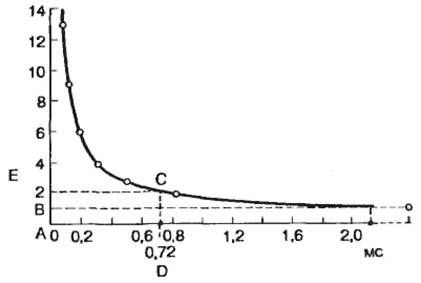
Скорость нарастания интенсивности раздражителя имеет существенное значение при определении возбудимой ткани, поэтому чаще всего используют импульсы прямоугольной формы (импульс тока прямоугольной формы имеет максимальную крутизну нарастания). Замедление скорости изменения амплитуды раздражителя приводит к тому, что происходит инактивация натриевых каналов вследствие постепенной деполяризации клеточной мембраны, а следовательно, к падению возбудимости. Увеличение силы стимула до порогового значения приводит к генерации потенциала действия (рис. 2.14, В). Под анодом при действии сильного тока происходит изменение уровня критического потенциала, в противоположном направлении — вниз (рис. 2.14, Д). При этом уменьшается разность между критическим потенциалом и мембранным потенциалом, т. е. возбудимость под анодом при длительном действии тока повышается. Очевидно, что увеличение значения тока до пороговой величины приведет к тому, что возбуждение будет возникать под катодом при замыкании цепи. Следует подчеркнуть, что этот эффект может быть выявлен в случае продолжительного действия электрического тока. При действии достаточно сильного тока смещение критического потенциала под анодом может быть весьма существенным и достигать первоначального значения мембранного потенциала. Выключение тока приведет к тому, что гиперполяризация мембраны исчезнет, мембранный потенциал вернется к первоначальному значению, а это соответствует величине критического потенциала, т. е. возникает анодно-размыкательное возбуждение. Изменение возбудимости и возникновение возбуждения под катодом при замыкании и анодом при размыкании носит название закона полярного действия тока. Экспериментальное подтверждение этой зависимости впервые было получено Пфлюгером еще в прошлом веке. Как указывалось выше, существует определенное соотношение между временем действия раздражителя и его амплитудой. Эта зависимость в графическом выражении получила название кривой «сила—длительность» (рис. 2.15). Иногда по имени авторов ее называют кривой Гоорвега—Вейса—Лапика. На этой кривой видно, что уменьшение значения тока ниже определенной критической величины не приводит к возбуждению ткани независимо от продолжительности времени, в течение которого действует этот раздражитель, а минимальная величина тока, вызывающая возбуждение, получила название порога раздражения, или реобазы. Величина реобазы определяется разностью между критическим потенциалом и мембранным потенциалом покоя. С другой стороны, раздражитель должен действовать не меньше определенного времени. Уменьшение времени действия раздражителя ниже критического значения приводит к тому, что раздражитель любой интенсивности не оказывает эффекта. Для характеристики возбудимости ткани по времени ввели понятие порога времени — минимальное (полезное)время, в течение которого должен действовать раздражитель пороговой силы с тем, чтобы вызвать возбуждение (отрезок АС на рис. 2.15). Порог времени определяется емкостной и резистивной характеристикой клеточной мембраны, т. е. постоянной временя T=RC. В связи с тем что величина реобазы может изменяться, особенно в естественных условиях, и это может привести к значительной погрешности в определении порога времени, Лапик ввел понятие хронаксии для характеристики временных свойств клеточных мембран. Хронаксия — время, в течение которого должен действовать раздражитель удвоенной реобазы, чтобы вызвать возбуждение. Использование этого критерия позволяет точно измерить временные характеристики возбудимых структур, поскольку измерение происходит на крутом изгибе гиперболы (отрезок AD на рис. 2.15). Хронаксиметрия используется при оценке функционального состояния нервно-мышечной системы у человека. При ее органических поражениях величина хронаксии и реобазы нервов и мышц значительно возрастает. Таким образом, при оценке степени возбудимости возбудимых структур используют количественные характеристики раздражителя — амплитуду, продолжительность действия, скорость нарастания амплитуды. Следовательно, количественная оценка физиологических свойств возбудимой ткани производится опосредованно по характеристикам раздражителя.
Переменный ток. Эффективность действия переменного тока определяется не только амплитудой, продолжительностью воздействия, но и частотой. При этом низкочастотный переменный ток, например частотой 50 Гц (сетевой), представляет наибольшую опасность при прохождении через область сердца. В первую очередь это обусловлено тем, что при низких частотах возможно попадание очередного стимула в фазу повышенной уязвимости миокарда (см. главу 7) и возникновение фибрилляции желудочков сердца. Действие тока частотой выше 10 кГц представляет меньшую опасность, поскольку длительность полупериода составляет 0,05 мс. При такой длительности импульса мембрана клеток вследствие своих емкостных свойств не успевает деполяризоваться до критического уровня. Токи большей частоты вызывают, как правило, тепловой эффект.
  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|