|
|
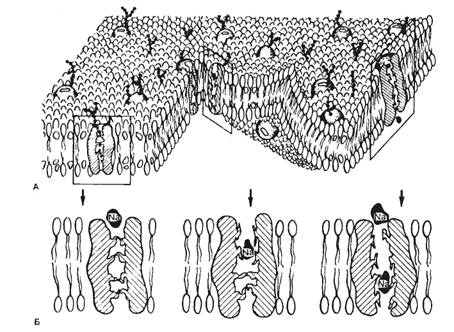
Строение и основные свойства клеточных мембран и ионных каналовСогласно современным представлениям, биологические мембраны образуют наружную оболочку всех животных клеток и формируют многочисленные внутриклеточные органеллы. Наиболее характерным структурным признаком является то, что мембраны всегда образуют замкнутые пространства, и такая микроструктурная организация мембран позволяет им выполнять важнейшие функции. Строение и функции клеточных мембран. 1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам. 2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»). 3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4.Высвобождение нейромедиаторов в синаптических окончаниях. Современными методами электронной микроскопии была определена толщина клеточных мембран (6—12 нм). Химический анализ показал, что мембраны в основном состоят из липидов и белков, количество которых неодинаково у разных типов клеток. Сложность изучения молекулярных механизмов функционирования клеточных мембран обусловлена тем, что при выделении и очистке клеточных мембран нарушается их нормальное функционирование. В настоящее время можно говорить о нескольких видах моделей клеточной мембраны, среди которых наибольшее распространение получила жидкостно-мозаичная модель. Согласно этой модели, мембрана представлена бислоем фосфолипидных молекул, ориентированных таким образом, что гидрофобные концы молекул находятся внутри бислоя, а гидрофильные направлены в водную фазу (рис. 2.1). Такая структура идеально подходит для образования раздела двух фаз: вне- и внутриклеточной.
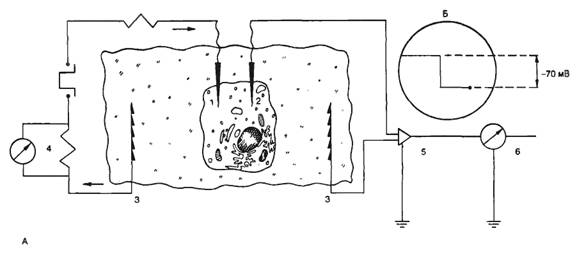
В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения. Здесь описана только общая схема строения клеточной мембраны и для других типов клеточных мембран возможны значительные различия. Электрические характеристики мембран. Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость. Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов. Кроме того, емкостные свойства клеточных мембран являются одной из причин, определяющих временные характеристики электрических процессов, протекающихщих на клеточных мембранах. Проводимость (g) — величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов. Через фосфолипидный бислой могут диффундировать различные вещества, причем степень проницаемости (Р), т. е. способность клеточной мембраны пропускать эти вещества, зависит от разности концентраций диффундирующего вещества по обе стороны мембраны, его растворимости в липидах и свойств клеточной мембраны. Скорость диффузии для заряженных ионов в условиях постоянного поля в мембране определяется подвижностью ионов, толщиной мембраны, распределением ионов в мембране. Для неэлектролитов проницаемость мембраны не влияет на ее проводимость, поскольку неэлектролиты не несут зарядов, т. е. не могут переносить электрический ток. Проводимость мембраны является мерой ее ионной проницаемости. Увеличение проводимости свидетельствует об увеличении количества ионов, проходящих через мембрану. Строение и функции ионных каналов. Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал (диаметр 0,5—0,7 нм). Расчеты показывают, что суммарная площадь каналов занимает незначительную часть поверхности клеточной мембраны. Функцию ионных каналов изучают различными способами. Наиболее распространенным является метод фиксации напряжения, или «voltage-clamp» (рис. 2.2). Сущность метода заключается в том, что с помощью специальных электронных систем в процессе опыта изменяют и фиксируют на определенном уровне мембранный потенциал. При этом измеряют величину ионного тока, протекающего через мембрану. Если разность потенциалов постоянна, то в соответствии с законом Ома величина тока пропорциональна проводимости ионных каналов. В ответ на ступенчатую деполяризацию открываются те или иные каналы, соответствующие ионы входят в клетку по электрохимическому градиенту, т. е. возникает ионный ток, который деполяризует клетку. Это изменение регистрируется с помощью управляющего усилителя и через мембрану пропускается электрический ток, равный по величине, но противоположный по направлению мембранному ионному току. При этом трансмембранная разность потенциалов не изменяется. Совместное использование метода фиксации потенциала и специфических блокаторов ионных каналов привело к открытию различных типов ионных каналов в клеточной мембране.
В настоящее время установлены многие типы каналов для различных ионов (табл. 2.1). Одни из них весьма специфичны, вторые, кроме основного иона, могут пропускать и другие ионы. Изучение функции отдельных каналов возможно методом локальной фиксации потенциала «path-clamp»; рис. 2.3, А). Стеклянный микроэлектрод (микропипетка) заполняют солевым раствором, прижимают к поверхности мембраны и создают небольшое разрежение. При этом часть мембраны подсасывается к микроэлектроду. Если в зоне присасывания оказывается ионный канал, то регистрируют активность одиночного канала. Система раздражения и регистрации активности канала мало отличается от системы фиксации напряжения. Таблица 2.1. Важнейшие ионные каналы и ионные токи возбудимых клеток
Примечание. ТЭА — тетраэтиламмоний; ТТХ — тетродотоксин. Ток через одиночный ионный канал имеет прямоугольную форму и одинаков по амплитуде для каналов различных типов (рис. 2.3, Б). Длительность пребывания канала в открытом состоянии имеет вероятностный характер, но зависит от величины мембранного потенциала. Суммарный ионный ток определяется вероятностью нахождения в открытом состоянии в каждый конкретный период времени определенного числа каналов (рис. 2.3, В).
Наружная часть канала сравнительно доступна для изучения, исследование внутренней части представляет значительные трудности. П. Г. Костюком был разработан метод внутриклеточного диализа, который позволяет изучать функцию входных и выходных структур ионных каналов без применения микроэлектродов. Оказалось, что часть ионного канала, открытая во внеклеточное пространство, по своим функциональным свойствам отличается от части канала, обращенной во внутриклеточную среду. Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость. Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. Канал неоднороден по своим функциональным характеристикам, особенно это касается белковых структур, находящихся у входа в канал и у его выхода (так называемые воротные механизмы).
Рассмотрим принцип работы ионных каналов на примере натриевого канала. Полагают, что в состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация) (рис. 2.4). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула. Работа натриевых каналов определяется величиной мембранного потенциала в соответствии с определенными законами вероятности. Рассчитано, что активированный натриевый канал пропускает всего 6000 ионов за 1 мс. При этом весьма существенный натриевый ток, который проходит через мембраны во время возбуждения, представляет собой сумму тысяч одиночных токов. При генерации одиночного потенциала действия в толстом нервном волокне изменение концентрации ионов Na+ во внутренней среде составляет всего 1/100000 от внутреннего содержания ионов Na гигантского аксона кальмара. Однако для тонких нервных волокон это изменение концентрации может быть весьма существенным. Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+, причем существуют разновидности каналов для этих ионов (см. табл. 2.1). Ходжкин и Хаксли сформулировали принцип «независимости» каналов, согласно которому потоки натрия и калия через мембрану независимы друг от друга. Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя. Особый интерес представляют кальциевые каналы. Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли «мессенджера», или вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током. Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца. Электрогенез кардиомиоцитов рассматривается в главе 7. Электрофизиологические характеристики клеточных мембран исследуют с помощью специальных методов. Методы изучения возбудимых клеток Электрические явления, которые возникают в возбудимых тканях, обусловлены электрическими свойствами клеточных мембран. Поэтому необходимо остановиться на методических подходах современной физиологии возбудимых тканей, используемых при исследовании электрических характеристик клеточных мембран. Любая физиологическая установка, предназначенная для изучения возбудимых клеток и тканей, должна содержать следующие основные элементы: 1) электроды для регистрации и стимуляции; 2) усилители биоэлектрических сигналов; 3) регистратор; 4) стимулятор; 5) систему для обработки физиологической информации. В зависимости от задач исследования обычно требуется дополнительное оборудование. Поскольку в современной медицине широко используются методы электрофизиологического исследования и воздействия электрическим током, необходимо кратко познакомиться с основными методическими приемами. При работе на изолированных органах, тканях и отдельных клетках применяют специальные камеры и растворы определенного состава, например Рингера-Локка, Тироде, Хэнкса, позволяющие в течение длительного времени поддерживать нормальную жизнедеятельность биологического объекта. Во время эксперимента раствор должен быть насыщен кислородом и иметь соответствующую температуру (для хладнокровных животных +20°С, для теплокровных +37°С). В процессе эксперимента необходимо использовать проточные камеры для непрерывного обновления раствора, в котором находится биологический объект. При электрофизиологических исследованиях используют различные типы электродов, детальное описание которых можно найти в соответствующих руководствах. В то же время есть определенные требования ко всем без исключения электродным системам. Электроды, которые используют в эксперименте, должны оказывать минимальное влияние на объект исследования, т. е. они должны только передавать информацию от объекта или на объект. Если в электрофизиологическом эксперименте исследуют собственно процесс возбуждения, то необходимо применять два электрода с различной величиной площади контактной поверхности (желательно в соотношении не менее 1:100), при этом электрод меньшей площади называют активным, или референтным, большей площади — пассивным, или индифферентным. При исследовании процесса распространения возбуждения необходимо использовать два активиых электрода с одинаковой площадью контактных поверхностей, устанавливаемых на возбудимой ткани на некотором расстоянии друг от друга, и индифферентный электрод, который устанавливается в отдалении. В первом случае говорят о моно-(уни-) полярном способе отведения потенциала (раздражении), во втором — о биполярном способе. Необходимо подчеркнуть, что термин «униполярный» способ весьма условен, поскольку всегда регистрируется разность потенциалов, а не абсолютное значение потенциала. Поскольку работа с биологическим объектом подразумевает контакт электрода с жидкостью, содержащейся в биологическом объекте, высока вероятность возникновения контактных поляризационных потенциалов, которые могут существенно исказить результаты исследования. Чтобы избежать возможных искажений в электрофизиологических экспериментах, как правило, используют специальные слабополяризующиеся электроды, например хлорсеребряные или каломельные, имеющие незначительный поляризационный потенциал. При исследовании электрофизиологических характеристик отдельных клеток используют стеклянные микроэлектроды. Они представляют собой микропипетку с диаметром кончика менее 0,5 мкм, заполненные ЗМ раствором хлорида калия. В электрофизиологических экспериментах применяют самые различные усилители биологических сигналов, позволяющие измерять минимальные изменения тока (до 10 А) и напряжения (до 10 -7 В) В связи с тем что регистрируемые сигналы могут иметь высокую скорость нарастания переднего фронта, усилители должны иметь достаточно широкую полосу пропускания (сотни кГц). Наибольшие требования предъявляются ко входным каскадам усилителей, которые должны быть согласованы с внутренним сопротивлением измерительного электрода, причем наибольшие трудности экспериментатор встречает при использовании микроэлектродов для регистрации быстрых изменений тока или потенциала, поскольку микроэлектроды могут иметь очень высокое внутреннее сопротивление (до 150 мОм). Стимулвторы, регистраторы, системы управления экспериментом и обработки физиологической информации еще более разнообразны и их описание можно найти в специальной литературе.
На рис. 2.5, А показана схема простейшей установки для измерения трансмембранной разности потенциалов и изучения реакций возбудимой мембраны при ее электрической стимуляции. Исследуемый биообъект (клетка, кусочек ткани) помещен в камеру, содержащую солевой раствор и электрод сравнения. Если измерительный электрод также находится в растворе, то разность потенциалов между ним и электродом сравнения стремится к нулю. В момент проникновения микроэлектрода внутрь клетки регистрируют отрицательный потенциал относительно внешней среды (рис. 2.5, Б). Перемещение кончика микроэлектрода внутри клетки не приводит к изменению измеряемой разности потенциалов, если электрод не повредил клетку. У покоящейся клетки с нормальным метаболизмом и стабильными условиями внешней и внутренней среды постоянная разность потенциалов будет регистрироваться неопределенно долго. Эта постоянная разность потенциалов называется потенциалом покоя, или мембранным потенциалом покоя. При этом потенциал внеклеточной среды принимается равным нулю. Величина потенциала покоя неодинакова у различных типов клеток и колеблется обычно от -70 до -95 мВ. В том случае, если в клетку введен второй, стимулирующий микроэлектрод, можно исследовать реакцию возбудимой мембраны на действие электрического тока. Если стимулирующий электрод электроотрицателен по отношению к внутренней среде клетки, то говорят о входящем токе, при этом общая трансмембранная разность потенциалов увеличивается, т. е. происходит гиперполяризация клеточной мембраны. Напротив, если стимулирующий электрод электроположителен по отношению к внутренней среде клетки, то говорят о выходящем токе, при этом общая трансмембранная разность потенциалов уменьшается, т. е. происходит деполяризация клеточной мембраны (рис. 2.6). Как правило, при действии гиперполяризующего тока потенциал мембраны изменяется в соответствии с законом Ома. При этом изменение потенциала не зависит от молекулярных процессов в мембране, поэтому говорят, что изменяются пассивные электрические свойства мембраны. При действии деполяризующего тока потенциал мембраны не подчиняется закону Ома, что связано с изменением функциональных характеристик ионных каналов клеточной мембраны. Если деполяризация клеточной мембраны достигает так называемого критического уровня, происходит активация ионных каналов клеточной мембраны и возникает потенциал действия. Критический потенциал (Eкp) — уровень мембранного потенциала, при котором начинается генерация потенциала действия. Потенциал действия (ПД, спайк, импульс) — быстрое колебание мембранного потенциала покоя в положительном направлении. В этом случае мембрана реагирует активно, поскольку изменение трансмембранной разности потенциалов обусловлено изменением функциональных свойств ионных каналов. Детальный анализ процессов, протекающих в мембранах возбудимых клеток, был проведен Ходжкиным, Хаксли и Катцем в опытах на гигантском аксоне кальмара и привел к созданию современной теории происхождения потенциала покоя и потенциала действия. Потенциал покоя Схема опыта Ходжкина—Хаксли приведена на рис. 2.7. В аксон кальмара диаметром около 1 мм, помещенный в морскую воду, вводили активный электрод, второй электрод (электрод сравнения) находился в морской воде. В момент введения электрода внутрь аксона регистрировали скачок отрицательного потенциала, т. е. внутренняя среда аксона была заряжена отрицательно относительно внешней среды.
Как указывалось в разделе 2.1.2, электрический потенциал содержимого живых клеток принято измерять относительно потенциала внешней среды, который обычно принимают равным нулю. Поэтому считают синонимами такие понятия, как трансмембранная разность потенциалов в покое, потенциал покоя, мембранный потенциал. Обычно величина потенциала покоя колеблется от -70 до -95 мВ. Согласно концепции Ходжкина и Хаксли, величина потенциала покоя зависит от ряда факторов, в частности от селективной (избирательной) проницаемости клеточной мембраны для различных ионов; различной концентрации ионов цитоплазмы клетки и ионов окружающей среды (ионной асимметрии); работы механизмов активного транспорта ионов. Все эти факторы тесно связаны между собой и их разделение имеет определенную условность. Известно, что в невозбужденном состоянии клеточная мембрана высокопроницаема для ионов калия и малопроницаема для ионов натрия. Это было показано в опытах с использованием изотопов натрия и калия: спустя некоторое время после введения внутрь аксона радиоактивного калия его обнаруживали во внешней среде. Таким образом, происходит пассивный (по градиенту концентраций) выход ионов калия из аксона. Добавление радиоактивного натрия во внешнюю среду приводило к незначительному повышению его концентрации внутри аксона. Пассивный вход натрия внутрь аксона несколько уменьшает величину потенциала покоя. Установлено, что имеется разность концентраций ионов калия вне и внутри клетки, причем внутри клетки ионов калия примерно в 20—50 раз больше, чем вне клетки (табл. 2.2). Таблица 2.2. Концентрация ионов снаружи и внутри клетки, ммоль/л
Разность концентраций ионов калия вне и внутри клетки и высокая проницаемость клеточной мембраны для ионов калия обеспечивают диффузионный ток этих ионов из клетки наружу и накопление избытка положительных ионов К+ на наружной стороне клеточной мембраны, что противодействует дальнейшему выходу ионов К+ из клетки. Диффузионный ток ионов калия существует до тех пор, пока стремление их двигаться по концентрационному градиенту не уравновесится разностью потенциалов на мембране. Эта разность потенциалов называется калиевым равновесным потенциалом. Равновесный потенциал (для соответствующего иона, Ек) — разность потенциалов между внутренней средой клетки и внеклеточной жидкостью, при которой вход и выход иона уравновешен (химическая разность потенциалов равна электрической). Важно подчеркнуть следующие два момента: 1) состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов (по сравнению с их общим содержанием); калиевый равновесный потенциал всегда больше (по абсолютному значению) реального потенциала покоя, поскольку мембрана в покое не является идеальным изолятором, в частности имеется небольшая утечка ионов Na+. Сопоставление теоретических расчетов с использованием уравнений постоянного поля Д. Голдмана, формулы Нернста показали хорошее совпадение с экспериментальными данными при изменении вне- и внутриклеточной концентрации К+ (рис. 2.8). Трансмембранная диффузионная разность потенциалов рассчитывается по формуле Нернста: Ek=(RT/ZF)ln(Ko/Ki) где Ек — равновесный потенциал, R — газовая постоянная, Т — абсолютная температура, Z — валентность нона, F — постоянная Фарадея, Ко и Ki — концентрации ионов К+ вне и внутри клетки соответственно.
Величина мембранного потенциала для значений концентрации ионов К+, приведенных в табл. 2.2, при температуре +20 °С составит примерно —60 мВ. Поскольку концентрация ионов К+ вне клетки меньше, чем внутри, Ек будет отрицательным. В состоянии покоя клеточная мембрана высокопроницаема не только для ионов К+. У мышечных волокон мембрана высокопроницаема для ионов СГ. В клетках с высокой проницаемостью для ионов Сl-, как правило, оба иона (Сl- и К+) практически в одинаковой степени участвуют в создании потенциала покоя. Известно, что в любой точке электролита количество анионов всегда соответствует количеству катионов (принцип электронейтральности), поэтому внутренняя среда клетки в любой точке электронейтральна. Действительно, в опытах Ходжкина, Хаксли и Катца перемещение электрода внутри аксона не выявило различие в трансмембранной разности потенциалов. Поскольку мембраны живых клеток в той или иной степени проницаемы для всех ионов, совершенно очевидно, что без специальных механизмов невозможно поддерживать постоянную разность концентрации ионов (ионную асимметрию). В клеточных мембранах существуют специальные системы активного транспорта, работающие с затратой энергии и перемещающие ионы против градиента концентраций. Экспериментальным доказательством существования механизмов активного транспорта служат результаты опытов, в которых активность АТФазы подавляли различными способами, например сердечным гликозидом оуабаином. При этом происходило выравнивание концентраций ионов К+ вне и внутри клетки и мембранный потенциал уменьшался до нуля. Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (рис. 2.9). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается с 3 находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с 2 находящимися вне клетки ионами К+, которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой схеме приводит к следующим результатам. 1. Поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится, активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе. Однако величина вклада электрогенного насоса в общее значение потенциала покоя обычно невелика и составляет несколько милливольт. 2. Поддерживается низкая концентрация ионов натрия внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки. 3. Поддерживая стабильный концентрационный градиент Na+, натрий-калиевый насос способствует сопряженному транспорту аминокислот и сахаров через клеточную мембрану. Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой проводимостью клеточной мембраны в состоянии покоя для ионов К+ (для мышечных клеток и ионов Сl-), ионной асимметрией концентраций для ионов К+ (для мышечных клеток и для ионов Cl-), работой систем активного транспорта, которые создают и поддерживают ионную асимметрию.
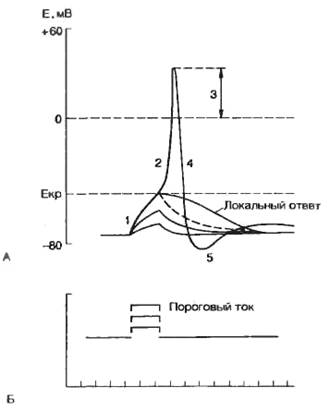
2.1.4. Потенциал действия Емкость мембраны и работа метаболических ионных насосов приводят к накоплению потенциальной электрической энергии на клеточной мембране в форме потенциала покоя. Эта энергия может освобождаться в виде специфических электрических сигналов (потенциала действия), характерных для возбудимых тканей: нервной, мышечной, некоторых рецепторных и секреторных клеток. Под потенциалом действия понимают быстрое колебание потенциала покоя, сопровождающееся, как правило, перезарядкой мембраны. Форма потенциала действия аксона и терминология, используемая для описания потенциала действия, приведены на рис. 2.10. Для правильного понимания процессов, происходящих при генерации потенциала действия, используем схему опыта, приведенную на рис. 2.5. Если через стимулирующий электрод подавать короткие толчки гиперполяризующего тока, то можно зарегистрировать увеличение мембранного потенциала, пропорциональное амплитуде подаваемого тока; при этом мембрана проявляет свои емкостные свойства — замедленное нарастание и снижение мембранного потенциала (см. рис. 2.6). Ситуация будет изменяться, если через стимулирующий электрод подавать короткие толчки деполяризующего тока. При небольшой (подпороговой) величине деполяризующего тока мембрана ответит пассивной деполяризацией и проявит емкостные свойства. Подпороговое пассивное поведение клеточной мембраны называется электротоническим, илиэлектротоном. Увеличение деполяризующего тока приведет к появлению активной реакции клеточной мембраны в форме повышения натриевой проводимости (gNa+). При этом проводимость клеточной мембраны не будет подчиняться закону Ома. Отклонение от пассивного поведения проявляется обычно при 50—80% значении порогового тока. Активные подпороговые изменения мембранного потенциала называются локальным ответом.
Смещение мембранного потенциала до критического уровня приводит к генерации потенциала действия. Минимальное значение тока, необходимого для достижения критического потенциала, называют пороговым током. Следует подчеркнуть, что не существует абсолютных значений величины порогового тока и критического уровня потенциала, поскольку эти параметры зависят от электрических характеристик мембраны и ионного состава окружающей внешней среды, а также от параметров стимула. Зависимость между величиной стимулирующего тока и временем его действия рассматривается в разделе 2.1.5. В опытах Ходжкина и Хаксли был обнаружен, на первый взгляд, удивительный эффект. Во время генерации потенциала действия мембранный потенциал уменьшался не просто до нуля, как следовало бы из уравнения Нернста, но изменил свой знак на противоположный. Анализ ионной природы потенциала действия, проведенный первоначально Ходжкиным, Хаксли и Катцем, позволил установить, что фронт нарастания потенциала действия и перезарядка мембраны (овершут) обусловлены движением ионов натрия внутрь клетки. Как уже указывалось выше, натриевые каналы оказались электроуправляемыми. Деполяризующий толчок тока приводит к активации натриевых каналов и увеличению натриевого тока. Это обеспечивает локальный ответ. Смещение мембранного потенциала до критического уровня приводит к стремительной деполяризации клеточной мембраны и обеспечивает фронт нарастания потенциала действия. Если удалить ион Na+ из внешней среды, то потенциал действия не возникает. Аналогичный эффект удавалось получить при добавлении в перфузионный раствор ТТХ (тетродотоксин) — специфического блокатора натриевых каналов (см. табл. 2.1). При использовании метода «voltage-clamp» было показано, что в ответ на действие деполяризующего тока через мембрану протекает кратковременный (1—2 мс) входящий ток, который сменяется через некоторое время выходящим током (рис. 2.11). При замене ионов натрия на другие ионы и вещества, например холин, удалось показать, что входящий ток обеспечивается натриевым током, т. е. в ответ на деполяризующий стимул происходит повышение натриевой проводимости (gNa+). Таким образом, развитие фазы деполяризации потенциала действия обусловлено повышением натриевой проводимости. Критический потенциал определяет уровень максимальной активации натриевых каналов. Если смещение мембранного потенциала достигает значения критического уровняпотенциала, то процесс поступления ионов Na+ в клетку лавинообразно нарастает. Система начинает работать по принципу положительной обратной связи, т. е. возникает регенеративная (самоусиливающаяся) деполяризация. Перезарядка мембраны, или овершут, весьма характерна для большинства возбудимых клеток. Амплитуда овершута характеризует состояние мембраны и зависит от состава вне- и внутриклеточной среды. На высоте овершута потенциал действия приближается к равновесному натриевому потенциалу, поэтому происходит изменение знака заряда на мембране.
Экспериментально было показано, что амплитуда потенциала действия практически не зависит от силы стимула, если он превышает пороговую величину. Поэтому принято говорить, что потенциал действия подчиняется закону "в<   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|