|
|
Гетерогенность модулирующей системы мозгаОдно время считалось, что, несмотря на большое разнообразие физиологических реакций (ЭЭГ, КГР, ЭКГ, ЭМГ и др.), которые обычно используются для оценки ФС, все они в одинаковой мере связаны с активацией неспецифической системы мозга и что по динамике любой из них можно предсказать изменение всех остальных реакций. Такой подход основывался на появившейся в 1951 г. концепции Д. Линдсли о единстве и синергизме восходящих и нисходящих влияний от неспецифической системы мозга, обусловливающих параллелизм всех ЭЭГ-х, вегетативных и моторных реакций активации. Однако позже были получены данные о низких коэффициентах корреляции между различными показателями активации, а также о диссоциации ЭЭГ и поведенческого пробуждения. Введение собаке атропина вызывает ЭЭГ медленного сна, в то время как животное поведенчески продолжает бодрствовать. С помощью физосигмина можно вызвать активированную ЭЭГ, тогда как животное будет находиться в состоянии дремоты. Многочисленные данные о разнонаправленном изменении различных показателей активации (в том числе ЭЭГ и ЧСС) были обобщены Дж. Лейси в его концепции «дирекционной фракционности активации». По Лейси, существует не единая система неспецифической активации, а несколько субсистем, выражением которых являются вегетативные, моторные и ЭЭГ-реакции. П.К. Анохин принадлежит концепция «специфичности неспецифической активации». Он утверждает, что каждый тип мотивации обеспечивается возбуждением собственной неспецифической активирующей системы, обладающей особой химической специфичностью. В опытах на животных он выделил и описал различные ЭЭГ паттерны для оборонительной, пищевой и ориентировочной мотивации и, используя различные фармакологические вещества, продемонстрировал возможность избирательной блокады каждого из этих состояний. Известна точка зрения A.A. Роутенберга, выделяющего две системы активации: лимбическую систему и ретикулярную формацию ствола мозга с их относительным антагонизмом. Активирующая ретикулярная формация обеспечивает энергетическую базу исполнения реакций и имеет тенденцию участвовать в оборонительном поведении, тогда как лимбическая система занята преимущественно активацией вегетативных процессов и связана с подготовительными фазами поведения. С помощью факторного анализа ЭЭГ затылка и лба В.М. Русаловым выделено четыре общемозговых интегральных ЭЭГ-фактора: 1) фактор энергии медленных волн; 2) фактор частоты медленных ритмов; 3) фактор активности бета-2; 4) фактор пространственно-временной сопряженности ЭЭГ. Факторы рассматриваются как отражение независимых аспектов регуляции уровня неспецифической активации, за которыми стоит функционирование раздельных и относительно самостоятельных систем активации мозга. В свете этих представлений функциональное состояние является результатом взаимодействия или баланса отдельных субсистем активации, т. е. ФС — явление системное, которое требует системного подхода. Серьезное подтверждение существования субсистем активации, взаимодействие которых определяет функциональное состояние человека, получено в психофизических экспериментах. В исследованиях Р. Тейера испытуемые ранжировали по 4-балльной системе собственные состояния (или «субъективную активацию») в соответствии с предложенным списком. Это был перечень прилагательных для различных состояний, укладывающихся в шкалу от напряженного бодрствования до спокойного состояния. Все эмоциональные состояния намеренно исключались. Методом факторного анализа было выделено два генеральных фактора регуляции ФС, взаимодействие которых и определяет особенности каждого состояния. Фактор I был определен как «общая активация и деактивация-сон» или шкала переживаний от бодрствования до сна. На одном полюсе — ощущение мощи, энергии, живости, на другом — ощущение сонливости, усталости. Фактор II («высокая активация и общая деактивация») представлял шкалу переживаний от напряженности до безмятежности и спокойствия. Изменение субъективной активации по шкале бодрствования хорошо коррелировало с изменением температуры тела в цикле бодрствование—сон, и физические упражнения увеличивали уровень субъективной активации по этой шкале. Усиление тревожности сопровождается ростом субъективной эмоциональной активации и падением субъективной активации по шкале бодрствования. Реципрокность изменений двух активации выявилась и в связи с циркадным ритмом. Две системы субъективной активации методом семантического дифференциала выделены В.И. Викторовым. Показано их различное взаимоотношение с успешностью обучения в вузе. Фактор А, охватывающий континуум состояний от спокойствия до волевого усилия, связан с успешностью обучения куполообразной зависимостью, тогда как фактор Р, соответствующий шкале переживаний от печали до радости, и успешность обучения связаны отрицательной линейной зависимостью. Несколько субсистем активации можно выделить и в семантическом пространстве «аффективных значений», полученных Ч. Осгудом методом семантического дифференциала. Ведущие факторы пространства Ч. Осгуда интерпретированы как оценка (1), сила (потенция) (2) и активность (3), где фактор активности соответствует шкале эмоциональности, фактор силы — шкале бодрствования, по Р. Тейеру, а фактор оценки — шкале приятных и неприятных переживаний. Существенной особенностью пространства аффективных значений Ч. Осгуда является его универсальность, т. е. независимость от выборки обследуемых, различающихся культурой, возрастом, уровнем интеллектуального развития, языком. Это указывает на то, что в пространстве аффективных значений, по-видимому, находят отражение некоторые общие физиологические закономерности работы мозга, имеющие отношение к базальным механизмам регуляции состояний человека. Подтверждение существования двух субсистем активации: эмоциональной и неэмоциональной — получено нами при построении семантических пространств состояний методом многомерного шкалирования (по алгоритму Торгерсона). Испытуемые ранжировали степень различия пар слов из списка, который включал как названия эмоций, так и состояний с минимальной эмоциональной компонентой (например, спокойствие, растерянность, уверенность и др.). Семантическое пространство состояний у всех оказалось трехмерным. Его оси ортогональны и интерпретированы как: 1) шкала знаков состояний (положительные — отрицательные состояния); 2) шкала бодрствования или готовности к действию (неэмоциональная активация — НЭА); 3) шкала выраженности эмоционального тона (эмоциональная активация — ЭА). Индивидуальные семантические пространства состояний различались субъективной значимостью трех основных осей пространства. У лиц с низкой реактивностью (по тесту Я. Стреляу) и высокой экстраверсией (по Г. Айзенку) шкала бодрствования (или неэмоциональной активации) имела более высокий удельный вес по сравнению со шкалой эмоциональной активации. Представление о системном характере регуляции ФС получило подтверждение и при изучении специального класса регуляторов ФС — волновых генераторов электрической активности мозга. Известно, что модулирующая система распространяет два типа влияний: 1) тонические, когда изменяется средний уровень возбудимости нервной системы, и 2) ритмические, когда возбудимость нервных элементов меняется периодически. Сейчас накоплены многочисленные данные о том, что поток возбуждения, поступающий в мозг по сенсорным путям, квантуется его ритмической активностью (тета-, альфа-активностью, волнами ЭКГ и др.). Ритмической модуляции подвергаются также и исполнительные, двигательные системы. Время простой сенсомоторной реакции, вероятность появления произвольной двигательной реакции модулируются периодом альфа-волны у человека. Ухудшение и улучшение зрительного восприятия эмоционального слова по показателю КГР связаны с различными фазами альфа-волны. Скорость привыкания также зависит от фазы альфа-волны, с которой совпадает подача повторяющегося стимула. Двигательные инструментальные реакции крысы — нажим на педаль и ее отпускание — соответствуют разным противоположным фазам тета-волн. Высказано предположение, что ритмическая активность мозга осуществляет квантование и в системе памяти. На основе изучения нейронных механизмов ритмической активности мозга у животных и ЭЭГ у человека выделено два самостоятельных волновых генератора: генератор низкочастотного (7-8 Гц) и среднечастотного (9-10 Гц) альфа-ритма. Они были идентифицированы как две субсистемы активации: эмоциональной и неэмоциональной. Оба генератора обладают резонансными свойствами. Их активность представлена в УВП разными компонентами. Конкретное ФС человека в условиях бодрствования определяется балансом двух систем ритмической активности. При этом чем выше активность среднечастотного альфа-генератора, тем ниже уровень активности низкочастотного генератора, и наоборот. Для среднечастотного альфа-генератора найден ЭЭГ-показатель его активности в виде реакции усвоения ритма световых мельканий, совпадающего с его резонансной частотой. Чем больше доля высоких гармоник в составе реакции усвоения, тем выше уровень активности среднечастотного генератора. Показателем активности низкочастотного альфа-генератора может служить вертекс-потенциал. Чем больше его амплитуда, тем выше активность низкочастотного альфа-генератора. Оптимальность ФС для выполняемой работы связана с преобладанием активности среднечастотного альфа-генератора над низкочастотным. Взаимоотношение этих двух волновых генераторов или двух систем активации и обусловливает колоколообразную форму кривой, которой обычно описывают отношения эффективности деятельности и ФС. Падение эффективности выполняемой работы при высоких уровнях активации нервной системы детерминировано сдвигом баланса двух систем активации в направлении преобладания системы ЭА над НЭА.
Сон Длительное время господствовало представление о том, что во время сна, когда в течение нескольких часов в организме идут процессы восстановления, наблюдается общее снижение активности тела и мозга. Такое понимание соответствовало особенностям сонного поведения, когда человек или животное, приняв удобную позу, многие часы находились без движения, а также регистрируемой при этом ЭЭГ, в которой появлялись медленные волны. Позже представление о падении активности мозга во время сна было отвергнуто прежде всего благодаря открытию особой стадии сна, так называемого парадоксального или быстрого сна. Его открытие в 1953 г. связано с именами аспиранта Э. Азеринского и исследователя Н. Клейтмена, которые во время ночного сна у здоровых людей зарегистрировали быстрые движения глаз. Периоды таких движений в течение ночи появлялись 4-5 раз и, как в дальнейшем было показано У. Дементом и Н. Клейтменом, они совпадали с десинхронизацией в ЭЭГ. По данным М. Жуве, десинхронизация ЭЭГ во время сна сочетается с фазическими двигательными реакциями в вице подергивания конечностей, вибрис, хвоста. Таким образом, сон — это не просто углубление и распространение торможения. Это сложный процесс, имеющий свои стадии, каждая из которых сама по себе уникальна. Различие между ними не может быть описано количественно. При этом мозг во время сна характеризуется высоким уровнем активности и в некотором смысле даже более высоким, чем при спокойном бодрствовании (рис. 46). Существует несколько классификаций ЭЭГ-стадий сна человека. Наибольшее распространение получила классификация, предложенная У. Дементом и Н. Клейтменом. Для I стадии характерна ЭЭГ с низковольтной, быстрой активностью, иногда включающей короткие группы альфа-волн; во II стадии в ЭЭГ на уплощенном фоне появляются сонные веретена (12-14 в/с), билатеральные «острые волны» иК-комплекс; в III стадии — сонные веретена в сочетании с высоковольтными дельта-волнами; на IV стадии развивается наиболее глубокий медленный сон: в ЭЭГ дельта-волны без веретен. Переход от бодрствования ко сну — это особое состояние, которое осуществляется через ряд гипнотических фаз, выделенных и описанных И.П. Павловым Рис. 46. ЭЭГ бодрствования и электроэнцефалографические стадии сна у человека. Стрелками показано чередование быстрого и медленного сна
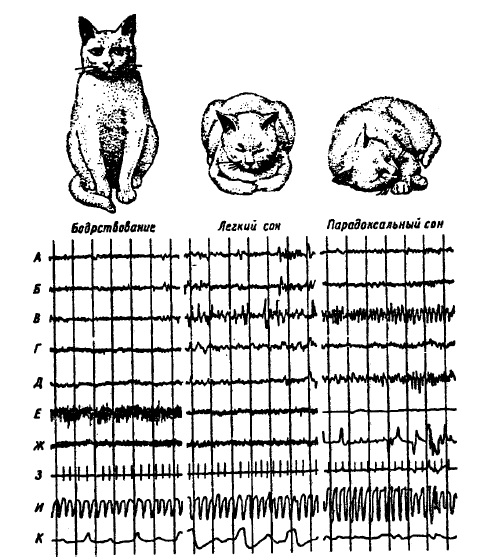
как уравнительная, парадоксальная, ультрапарадоксальная и наркотическая. Гипнотические фазы различают по величине условных рефлексов на сильные и слабые, положительные и отрицательные условные раздражители. У. Демент, Н. Клейтмен описали и V стадию сна как парадоксальный сон с ЭЭГ десинхронизированного типа и с появлением во время этой стадии энергичных и быстрых движений глаз (БДГ), которые иногда сопровождаются подергиванием конечностей, туловища. Качественное отличие V стадии сна от всех других позволило Л. Джонсону настаивать на классификации стадий сна, признающей только две его фазы: с БДГ и без БДГ. Три основные состояния мозга кошки: бодрствование, легкий и глубокий (парадоксальный) сон — показаны на рис. 47. Нормальная кошка примерно две трети времени проводит во сне. Он начинается с 25-минутного легкого сна, который сопровождается 6-7-минутным парадоксальным сном. Во время последнего ее мышцы расслаблены, ее трудно разбудить. Как медленный, так и быстрый сон характеризуются тоническими и фазическими явлениями. Наиболее существенное изменение во время медленного сна по сравнению с бодрствованием — возрастание порогов поведенческого пробуждения на электрическую стимуляцию ретикулярной формации среднего мозга, которые, по данным М. Жуве, увеличиваются на 30-40%. Медленный сон развивается при снижении тонической активности антигравитационной мускулатуры (начало атонии), но иногда уровень мышечного тонуса может оставаться таким же, как и во время бодрствования. На этом фоне могут наблюдаться фазические явления: клонические подергивания конечностей и шеи. У человека эти реакции сопровождаются возрастанием частоты дыхания и сужением Рис. 47. Бодрствование, легкий сон и парадоксальный сон у кошки. Рис. 47. Бодрствование, легкий сон и парадоксальный сон у кошки. Сверху вниз: электрическая активность: А — сенсомоторной коры, Б — эктосильвиевой коры, В — вентрального гиппокампа, Г — ретикулярной формации среднего мозга, Д — моста, Е — мышц шеи, Ж — движения глаз, 3 — ЭКГ, И — плетизмограмма, К — дыхание; вертикальные линии — время в секундах (по М. Жуве, 1967)
сосудов пальца. Увеличивается число спонтанно появляющихся электродермальных ответов (кратковременные изменения кожной проводимости), так называемое явление «вегетативного шторма». Во время MC в ЭЭГ-синхронизированные медленные волны, при этом у части нейронов частота спайковых разрядов в фоне падает, у другой растет. К фазическим явлениям ЭЭГ относят сонные веретена — серии колебаний частотой 11-16 в/с. Их появление совпадает с пачечными разрядами волокон пирамидного тракта и отрицательным сдвигом постоянного потенциала в спинном мозге на уровне мотонейронов (α-мотонейронов). Эти изменения рассматриваются О.Помпеано как причина клонических подергиваний конечностей. М. Жуве одновременно с веретенами наблюдал появление фазической мускульной активности шеи. На II стадии медленного сна сонное веретено часто сопровождается К-комплексом — серией медленных потенциалов большой амплитуды. За ним следуют реакции автономной нервной системы: возрастание частоты сердечных ударов и сосудистая реакция пальцев. Нередко к этим вегетативным реакциям добавляются общие движения тела, обычно с запаздыванием в среднем на 2,52 с. Данный комплекс фазических реакций возникает с определенной регулярностью, что свидетельствует о его эндогенном происхождении. Частота появления веретен увеличивается перед ПС. Наиболее типичным поведенческим проявлением парадоксального сна является полное расслабление мускулатуры тела, поддерживающей позу животного или человека, т. е. полная атония антигравитационной мускулатуры и исчезновение активности мышц шеи. По сравнению с медленным сном вегетативные изменения в ПС выражены более отчетливо. Так, у кошек резко снижается частота сердечных ударов и падает кровяное давление (см. рис. 47). У человека же в ПС частота сердечных ударов и давление растут. Дыхание и у человека, и у животных становится нерегулярным. Электродермальная активность заметно падает по сравнению с медленным сном. Парадоксальный сон отличается своеобразными фазическими явлениями — быстрыми движениями глаз. Они появляются в виде взрывов, включающих от 5 до 50 движений. Их частота 60-70 движений в/мин. Они не похожи на движения глаз при рассматривании картины. Но некоторые исследователи находят их сходство с движением глаз, возникающим при запоминании зрительных изображений. БДГ человека занимают около 10% времени всей стадии ПС и, как правило, возникают в комплексе с движениями тела, конечностей и с фазической вегетативной иррегулярностью. Развитию движения глазных яблок у животных нередко предшествует резкое усиление гаппокампального тета-ритма. Предполагают, что комплекс фазических явлений ПС — это ЭЭГ и соматические проявления эмоциональных реакций, возникающих на фоне эмоционального возбуждения в условиях сильной заторможенности спинального аппарата двигательных реакций. ЭЭГ парадоксального сна сходна с ЭЭГ бодрствования. Как правило, во время ПС наблюдается десинхронизация электрической активности мозга. Однако у человека, у которого альфа-ритм хорошо выражен, он также может регистрироваться и во время ПС. У животных же во время ПС обычно виден тета-ритм, особенно в лимбических структурах, при этом он более высокочастотен и регулярен по сравнению с бодрствованием (см. рис. 47). Регистрация нейронной активности в ретикулярной формации среднего мозга, заднего гипоталамуса, поясной извилине во время цикла бодрствование — сон показывает ее удивительное сходство во время ПС и бодрствования. Это позволяет некоторым исследователям говорить о ПС как об аналоге бодрствования с тем лишь различием, что при ПС возникает атония скелетных мышц и резко снижается активность сенсорных входов, тогда как процессы, происходящие в головном мозге, качественно сходны. По данным Т.Н. Ониани, следует выделять две стадии ПС: эмоциональную и неэмоциональную. У животных первая характеризуется тета-ритмом и быстрыми движениями глаз, вторая — ЭЭГ-десинхронизацией и отсутствием БДГ. Он связывает их соответственно с выражением эмоционально-мотивационного возбуждения и состояния удовлетворения потребности, которые за время ПС несколько раз сменяют друг друга. Эмоциональная и неэмоциональная стадии ПС рассматриваются как аналоги различных уровней бодрствования. В Варолиевом мосту, латеральном коленчатом теле таламуса и в зрительной коре регистрируются так называемые понто-геникуло-окципитальные спайки (ПГО). Это монофазные выбросы с такой же частотой, как и БДГ, т. е. 60-70 в/мин. Их появление на 0,6-1,5 мин опережает БДГ. Они являются первыми признаками ПС. Они первыми и исчезают за 1-2 мин до кортикальной активации и восстановления тонуса мышц шеи. Таким образом, наступление ПС характеризуется следующей последовательностью событий: сначала появляются ПГО, затем БДГ и с некоторым отставанием десинхронизация в ЭЭГ и исчезновение мышечного тонуса. Во время ПС пороги поведенческого пробуждения, вызываемого сенсорной стимуляцией, увеличиваются на 200-300% относительно бодрствования. При этом растет внутренняя активность мозга, что выражается в движениях глаз, конечностей, вегетативных реакциях при одновременном ухудшении проведения возбуждения по сенсорным путям. Естественный сон характеризуется циклической сменой медленного и парадоксального сна. Полный цикл, состоящий из смены медленного сна на быстрый с последующим возвращением к медленному, у человека занимает 60-90 мин. На ночной сон приходится 4-5 полных цикла. Сон всегда начинается с медленного сна (рис. 48). У нормального человека на быстрый сон в среднем приходится около 20% от общей продолжительности сна. Длительность фазы быстрого сна к утру увеличивается,.а медленного уменьшается, так что до 72% медленного сна расходуется в течение первой половины ночи. Фазические компоненты ПС более выражены в последних циклах сна. Структура сна у большинства млекопитающих сходна. Различия касаются в основном числа циклов смены MC на ПС. Так, у кошки в течение 5-часового периода развивается 10 циклов, тогда как у крысы количество циклов за это же время достигает 22. Такое
Р ис. 48. Типичное чередование фаз сна у взрослого молодого человека. Рис. 48. Типичное чередование фаз сна у взрослого молодого человека. Д — бодрствование, MC — медленный сон. Черной линией показаны периоды парадоксального сна (ПС) с характерной ЭЭГ-десинхронизацией и быстрыми движениями глаз (БДГ) (по Р. Томпсону, 1974)
быстрое чередование циклов возможно лишь за счет укорочения фаз MC и ПС. Парадоксальный сон имеется у всех млекопитающих, а также у птиц. У только что родившегося ребенка примерно половина всего сна приходится на ПС. И только с возрастом он уменьшается за счет появления MC. У недоношенных детей продолжительность ПС еще больше. Предполагают, что доминирование парадоксального сна у новорожденного способствует созреванию нервных элементов и формированию нервных связей, что достигается, в частности, за счет высокого уровня активности в ретикулярной системе (рис. 49). Хотя фазические явления ПС: ПГО и БДГ возникают почти одновременно, их происхождение связывают с разными структурами мозга. Быстрые движения глаз устраняются с разрушением вестибулярных ядер в продолговатом мозге (медиального и нисходящего ядра). При этом изолированные движения глаз
Рис. 49. Изменение соотношения длительности бодрствования (Б), медленного сна (MC) и парадоксального сна (ПС) с возрастом у человека (по Р. Томпсону, 1975)
сохраняются. Вместе с БДГ устраняется и весь комплекс реакций, который обычно им сопутствует: клонические подергивания конечностей, вегетативные реакции, фазическое расширение зрачка и др. Хотя БДГ и инициируются вестибулярными ядрами, однако их окончательное формирование зависит от двухолмия и РФ среднего мозга. Структура, от которой зависит генерация ПГО, — латеральная часть покрышки моста ростральнее вестибулярных ядер. Падение мышечного тонуса тела во время ПС связывают с сохранностью средней части моста. По данным Дж. Росси, М. Жуве, это дорзальная часть медиолатеральной покрышки моста, над зоной, ответственной за ПГО. Позже было показано, что возникновение атонии и ПГО зависит и от синего пятна. Разрушение структуры мозга, приводящее к устранению атонии мышц во время ПС, сопровождается странным поведенческим эффектом. С переходом от медленного сна к парадоксальному животное неожиданно встает, демонстрируя вегетативные и двигательные признаки страха, но не реагирует на сенсорные раздражения. Первые попытки объяснить механизмы развития сна разделили исследователей на сторонников «пассивной» и «активной» теорий сна. Сторонники «пассивной» теории связывали сон с деактивацией организма, которая может возникать в результате истощения или утомления нервных клеток самой РФ, или воздействия на нее каких-либо биохимических или гуморальных факторов, вырабатываемых во время бодрствования. Сторонники пассивных теорий не стремились объяснить сон, а лишь только то, почему невозможно бодрствование. Для них не существовало вопроса: «Почему человек заснул?», он был заменен вопросом: «Почему человек не способен бодрствовать?». Сторонники теории активного торможения признают необходимым воздействие одной структуры мозга на другую. С их точки зрения, деактивация РФ — вторичное явление и возникает вследствие тормозного влияния на РФ через кортикофугальные связи или со стороны тормозных структур мозга. И.П. Павлов выделял два механизма развития сна, которые, по существу, подтверждают правомерность позиций сторонников как пассивной, так и активной теории сна. С одной стороны, сон возникает как явление охранительного торможения, в результате сильного и длительного раздражения какого-либо отдельного участка коры больших полушарий. С другой стороны, сон возникает как результат внутреннего торможения, т. е. активного процесса формирования отрицательного условного рефлекса. Переход выработанного внутреннего торможения в сон экспериментально был многократно подтвержден. Таким образом, была выявлена условнорефлекторная природа сна и показано значение кортикофугальных влияний на его развитие. Во время бодрствования ретикулярная формация активирована и поддерживает тонус неокортекса, вызывая ЭЭГ-активацию и поведение бодрствования. Состояние сна развивается, когда возбуждается система ядер шва, по-видимому, в результате влияний из орбитофронтальной коры и преоптической области гипоталамуса, которая и тормозит ретикулярную формацию. Это позволяет неспецифическому таламусу проявить свои синхронизирующие свойства и вызвать в ЭЭГ коры больших полушарий медленные волны сна. Во время MC группа гигантских ретикулярных нейронов, которые находятся в реципрокных отношениях с нейронами синего пятна, периодически становится активной. Этому соответствует появление ЭЭГ-активации, движений глаз, ПГО, расслабление мышц шеи и другие признаки ПС. Структуры таламуса выполняют функцию «пейсмекера» для вызова ритмических потенциалов веретен во сне и альфа-ритма в бодрствовании. Активация ретикулярной формации блокирует синхронизацию ЭЭГ, вызываемую таламусом. Таламокортикальный механизм можно рассматривать как механизм внутреннего торможения, способного изменять активность мозга частично или глобально таким образом, что сенсорные, моторные и высшие функции мозга подавляются. Такой взгляд согласуется с представлением И.П.Павлова, что сон и внутреннее торможение — явления однородные и одно может переходить в другое. Возникновение синхронизированной активности в таламусе — начало цепочки реакций, приводящих к развитию медленного и парадоксального сна. Таким образом, сон — это активный процесс, который вызывает активацию одних структур и торможение других. В основе MC и ПС — различные системы интеграции структур мозга, предназначенные для выполнения различных функций. Чтобы понять функциональное назначение различных фаз сна, исследователи обратились к изучению психической деятельности человека во время сна. Одним из ее наиболее ярких проявлений являются сновидения. Одно время считалось, что сновидения видят не все. Обследование 5640 жителей Москвы, проведенное в 1974 г. под руководством A.B. Вейна, показало, что 48% опрошенных часто видят сны, причем у 19% сновидения носят устрашающий характер, 16% помнят цвет сновидений. Сновидения, как правило, носят зрительный характер. Реже встречаются сновидения с преобладанием слуховых и осязательных компонентов. Считают, что каждый человек в течение ночи видит 3-4 сна, но при пробуждении о них забывает.Была обнаружена связь сновидений с быстрыми движениями глаз. Так, у здоровых испытуемых представленность БДГ больше, чем у больных с нарушением сна, отчеты здоровых лиц о сновидениях также более ярки. Имеются данные, что частота БДГ связана со степенью личного участия в сновидениях и с яркостью снов. Отчет по сновидениям чаще возникает после пробуждения от ПС. Движения тела во время ПС связывают с окончанием сновидений и с переходом от одного сюжета к другому. Однако имеется ряд работ, в которых не подтверждается связь сновидений с БДГ. Поэтому исследователи высказывают разные точки зрения. Одни предполагают полную независимость распределения БДГ от содержания сновидений. Другие считают, что сновидения лишь модифицируют работу механизма, находящегося в стволе мозга и генерирующего БДГ. Однако многие склоняются к тому, что по интенсивности БДГ все же можно судить о яркости и эмоциональной насыщенности сновидений. Сновидения связаны с вегетативными изменениями во время ПС. По некоторым данным, приятные сны сочетаются с высокой частотой пульса. По другим, относительное учащение пульса в ПС по сравнению с MC положительно коррелирует с более содержательными отчетами о сновидениях, а при отсутствии такого учащения имеет место либо отрицание сновидений, либо забывание их содержания. Сначала казалось, что сновидения связаны только с ПС, так как у большинства людей после пробуждения от MC, как правило, не удавалось получить отчет о сновидениях. Однако более тщательное изучение показало, что в 64% пробуждений от MC человек рассказывает о психических переживаниях. Причем они скорее напоминают не сновидения, а мысли, рассуждения. Между сновидными переживаниями в MC и ПС существуют существенные различия. В MC во время сновидений зрительные картины менее четки, менее аффективны, менее длительны и более реальны. Тем не менее отчеты о сновидениях удалось получить не только после пробуждения от поверхностной (II) стадии сна, но и от глубокого сна (IV стадии). Во время медленного сна имеет место своеобразная психическая активность. На это указывают приступы сомнамбулизма, которые возникают только во время MC. Во время снохождения по одним данным в ЭЭГ возникают большие дельта-волны, по другим — альфа-ритм, который, однако, отличается тем, что он не подавляется световыми и другими раздражителями, не исчезает при открывании глаз и в этом отношении сходен с альфа-ритмом при гипнозе. Возникновение ночных кошмаров у детей и взрослых также связано с MC. Процессы, происходящие во время сна, имеют отношение к памяти. Хорошо известно, что заучивание материала перед сном помогает лучше его запомнить. Кроме того, если после заучивания материала следует 8-часовой сон, то его воспроизведение будет более успешным, нежели после 8 ч бодрствования. Особенно под влиянием сна улучшается запоминание бессмысленного материала. Запоминание улучшается главным образом после MC. Заученный материал лучше воспроизводится после первой половины ночи, чем после второй, когда преобладает ПС и почти отсутствует глубокий медленный сон. Однако позже было высказано предположение о важности ПС для запоминания. В основном это заключение основывалось на опытах с избирательной депривацией ПС, в которых было показано, что депривация ПС ведет к ухудшению формирования условных рефлексов у животных. Однако в лаборатории Т.Н. Ониани установили, что ухудшение обучения связано не с депривацией ПС, а со стрессом, которым сопровождается наиболее стандартная процедура депривации, когда кошку на длительное время помещали на маленькую платформу в бассейне, что не позволяло ей засыпать ПС (с атонией мышц). Если же депривацию ПС производить осторожным «подбуживанием» животного во время ПС, т. е. методом неэмоциональной и длительной (8-10 мин) депривации, то такая депривация не оказывает существенного влияния на обучение. Согласно Т.Н. Ониани, значение ПС для памяти состоит в том, что во время ПС происходит воспроизведение прошлого опыта, оживление следов долговременной памяти и тем самым задержка процесса забывания. Другая точка зрения на функцию ПС в отношении памяти сводится к тому, что во время ПС запоминается преимущественно биологически значимый материал. Еще относительно недавно интерес ко сну был прикован в связи с проблемой обучения во сне (гипнопедией). Первые результаты казались обнадеживающими. Однако использование полиграфической регистрации функционального состояния человека во время сеанса показало, что успешная гипнопедия требует поверхностного сна или даже пробуждения с альфа-ритмом. Чем глубже физиологический сон, тем меньше возможности для гипнопедии. т. е. этот метод нельзя считать полезным. Функциональную значимость фаз сна (MC и ПС) изучали также методами депривации сна. При длительном тотальном лишении сна в опытах Хайнемана до 116 ч наблюдались расстройства поведения, психических процессов, аффективной сферы, появление галлюцинаций, особенно зрительных. В первую восстановительную ночь преобладает глубокий медленный сон, тогда как после избирательной депривации ПС в восстановительную ночь обычно наблюдали удлинение ПС и увеличение БДГ. Первоначально полагали, что длительная депривация ПС приводит к изменениям поведения животных и человека, вызывая повышенное возбуждение, страхи, галлюцинации, нарушение моторных координаций и т. д. Однако затем эффект депривации оказался значительно менее сильным, чем при депривации только MC. Кроме того, он зависел от индивидуальных различий. У тех испытуемых, у которых возникали сновидения, несмотря на депривацию ПС, эта процедура не влияла на поведение и не вела к компенсаторному увеличению ПС в восстановительную ночь. Лица, у которых под влиянием депривации ПС возникали нарушения в поведении, испытывали страх, галлюцинации, во время процедуры многократно стремились к возобновлению ПС. По данным Т.Н. Ониани, ПС может быть заменен на бодрствование без большого ущерба для животного. Наиболее ранними теориями сна были гуморальные. Экспериментально были выделены некоторые химические вещества, которые циркулируют в крови и способны вызывать сон. М. Монье, Л. Гёзли выделили путем диализа из крови спящего кролика вещество, вызывающее сон (пептид-дельта сна или «дельта-фактор»). Другой гипногенный фактор («фактор сна»), лишенный видовой специфичности, был выделен Папенхаймером из ликвора коз, подвергнувшихся депривации сна. Сходный «фактор сна» был получен из ткани мозга животных на бойне, затем был обнаружен в моче человека. Анализ химической структуры этого пептида показал, что он относится к бактериальным (из материала оболочек бактерий) и накапливается в организме для дальнейшего использования в иммунной защите. Гипногенный эффект этих пептидов побочный. В начале 70-х гг. М. Монье проводил опыты на крысах, у которых длительное время вызывал поведение активного избегания, что сопровождалось стрессом. Затем из ткани мозга получил экстракт вещества мозга и выделил четыре различные фракции с гипногеяным эффектом («вещество сна»). Введение этих фракций через канюли в область таламуса вызывало увеличение MC и ПС. Согласно Я. Освальду, MC нужен для восстановления деятельности соматических органов, а функцией ПС является восстановление работоспособности клеток головного мозга. Только во время MC из гипоталамуса в кровь выбрасывается гормон роста, он участвует в биосинтезе белков в периферических тканях. Биосинтез белков и РНК нейронов интенсифицируется во время ПС. По Г. Лабори, MC связан с метаболической активностью нейроглии. Дж. Моруцци различает два типа восстановительных процессов в нервных клетках мозга. Быстрые восстановительные процессы, которые имеют место в нейронах, выполняющих функцию проведения и синаптической передачи нервных импульсов. Эти процессы завершаются в течение нескольких секунд и могут иметь место во время бодрствования без специального перерыва в активности нейронов. Сон не нужен для этих быстрых восстановительных процессов, которые, вероятно, доминируют в «шаблонных» синапсах, большей частью вовлекаемых во врожденную стереотипную активность. Медленные восстановительные процессы необходимы нейронам с синапсами, подверженными пластическим изменениям при обучении, восприятии и всех видах сознательной жизни, которые связаны с высшими функциями. Отсюда сон — это не период восстановления всего мозга, а только (или главным образом) период восстановления синапсов с пластическими свойствами. Большое распространение получило представление о мотивационных функциях ПС. Основываясь на новых данных, оно во многом созвучно положению 3. Фрей   Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|