
|
|
МЕЖДУНАРОДНАЯ КЛАССИФИКАЦИЯ ФЕРМЕНТОВСтр 1 из 8Следующая ⇒ Пояснительная записка
Курс биологической химии является теоретической основой любой пищевой технологии. Методическая разработка предназначена в помощь студентам заочной формы обучения для подготовки к экзамену и выполнения контрольной работы, а также студентам очного отделения для контроля самостоятельной работы. На современной этапе развития биологической химии достигнуты значительные успехи в области понимания молекулярных механизмов генетических процессов, изучения синтеза, структуры и функций белков и нуклеиновых кислот – главных молекул всего живого. В методической разработке рассматриваются биологически важные соединения - пептиды, белки, нуклеиновые кислоты, их строение, свойства, основные процессы с их участием и их механизмы. Разработка включает теоретический материал и методические рекомендации по решению упражнений и задач по следующим разделам программы дисциплины «Биологическая химия»: 1. Пептиды (строение, номенклатура, изомерия), белки. 2. Международная классификация ферментов. 3. Нуклеиновые кислоты (химические состав, строение). 4. Генетические процессы с участием нуклеиновых кислот и их закономерности (репликация ДНК, транскрипция ДНК, трансляция наследственной информации (биосинтез белка)). 5. Идентификация биологически важных соединений.
ПЕПТИДЫ, БЕЛКИ
Пептиды управляют жизненными функциями любого организма. Роль пептидов в процессах жизнедеятельности крайне многообразна. Многие из них служат гормонами (окситоцин, вазопрессин, гастрин, глюкагон, инсулин и др.), некоторые представлены сильнейшими ядами (яды змей, жаб, улиток, пауков, насекомых, высших грибов, микроорганизмов), мощными антибиотиками (пептиды микробов), ризилинг-факторами (способствуют синтезу и высвобождению гормонов), регуляторами клеточного деления, переносчиками молекул и ионов через биологические мембраны, регуляторами психической деятельности (способны вызывать радость и страх, гнев и удовольствие, могут ослаблять или усиливать боль, улучшать или ухудшать память), регуляторами физиологического состояния (могут влиять на чувство голода, работу сердца, сосудов и др.). В настоящее время насчитывается более 50 классов пептидов. Пептиды образуются в самых различных тканях человека. Интерес к ним объясним, так как их избыток или недостаток лежит в основе многих заболеваний. У здорового человека баланс пептидов поддерживается с высокой точностью. Значительное число природных пептидов синтезировано; кроме того, получены сотни аналогов, некоторые из которых обладают более сильным биологическим действием, нежели их природные предшественники. И те, и другие находят широкое практическое применение в медицинской практике, широко используются в пищевых технологиях в качестве вкусовых добавок – заменителей сахара (например, аспартам). Структурными элементами пептидов и белков являются остатки α-аминокислот. Пептидами называют соединения, построенные из небольшого числа молекул α-аминокислот, а системы, состоящие из множества соединенных между собой пептидных звеньев, называют полипептидами. Условно принято, что пептиды содержат от двух до 50 аминокислотных остатков, полипептиды – не более 100; если же их число превышает 100, то это белки. Остатки α-аминокислот во все перечисленных соединениях связаны между собой ковалентными пептидными связями (– CO – NH –). В состав белков входит 20 протеиногенных α-аминокислот (табл. 1). Протеиногенными называют α-аминокислоты, которые постоянно встречаются в природных белках, кодируются генетическим кодом и включаются белки в процессе их биосинтеза – трансляции.
Таблица 1 Протеиногенные α-аминокислоты
Все протеиногенные аминокислоты характеризуются общей структурной особенностью – наличием карбоксильной группы (– COOH) и аминогруппы (– NH2), которые связанны с одним и тем же атомом углерода, находящимся в α-положении по отношению к карбоксильной группе:
H2N – CH – COOH │ R Различаются же аминокислоты только боковыми цепями (R-группами), которые у разных аминокислот неодинаковы по структуре. Образование пептидов можно условно показать на примере межмолекулярного взаимодействия двух любых аминокислот друг с другом. При этом из двух молекул аминокислот образуется дипептид, из трех – трипептид и т.д. (для того, чтобы понять закономерность связывания остатков аминокислот в пептидах и белках, необходимо записать общий структурный блок аминокислот, а боковые цепи, связанные с α-углеродным атомом, убрать вниз). Межмолекулярное взаимодействие α-аминокислот осуществляется по типу реакции поликонденсации (образование пептидных связей (– CONH –) сопровождается выделением низкомолекулярного вещества – воды):
H2N–CH2–COOH+H NH–CH–COOH → H2N–CH2 –CO –NH –CH –COOH→ │ -H2O │ CH3 дипептид CH3
-H2O │ CH3 трипептид Исключение: в образовании пептидной связи у пролина (иминокислота) принимает участие α-иминогруппа, в этом случае пептидная связь будет – CO – N –:
| / \ -H2O | / \ CH3 H2C CH2 CH3 H2C CH2 \ / \ / CH2 CH2
Таким образом, характерной особенностью строения полипептидной цепи является наличие монотонно повторяющихся –NH–CH–CO– фрагмен- | тов, которые образую скелет, или остов молекулы. Полипептидная цепь является структурной основой белков. В действительности синтез пептидов и полипептидов из аминокислот – очень сложный процесс (в клетках они синтезируются в процессе трансляции на рибосомах). Протеиногенные аминокислоты способны включаться в пептиды, полипептиды и белки в процессе их биосинтеза в различных комбинациях и последовательностях (в соответствии с генетической информацией), в результате образуются пептиды, полипептиды и белки, обладающие совершенно разными свойствами и биологической активностью. Подсчитано, что из 20 аминокислот можно получить 2,4·1018 различных комбинаций, т.е. изомеров, но ведь это очень короткий полипептид! Сколько же может существовать аминокислотных последовательностей? Дипептид, состоящий их двух разных аминокислот – А и В, в зависимости от порядка их расположения, может иметь две изомерные формы: АВ и ВА. Трипептид, состоящий из трех различных аминокислот – А, В и С, может существовать в шести, отличающихся по последовательности, вариантах: АВС, АСВ, ВАС, ВСА, САВ, и СВА. Число возможных изомеров по последовательностям аминокислот, при условии отсутствия повторов, можно рассчитать по формуле «эн факториала»: n! = n (n–1)(n–2) и т.д., где n – число аминокислот. Например, трипептид из трех разных аминокислот может иметь 3! = 3 (3–1)(3–2) = 6 различных изомеров. Для полипептида из 10 различных аминокислот, ни одна из которых не повторяется в нем дважды, число изомеров составит 10! = 10 · 9 · 8 · 7 · 6 · 5 · 4 · 3 · 2 ·1, что дает в результате цифру равную 3628800. Сколько различных дипептидов может существовать при условии, что любая из двух аминокислот (А и В) может занимать любое из двух возможных положений, причем каждую аминокислоту можно использовать более одного раза? Дипептид может иметь следующие изомерные формы: АВ, ВА, АА, ВВ. Отсюда следует, что число возможных изомеров в этом случае можно рассчитать по формуле: Аn: где n – число возможных повторов, А – общее количество аминокислот, т.е. в приведенном примере число возможных изомеров дипептида будет: 22 = 4. Следовательно, 20 протеиногенных аминокислот при условии их повторов дают достаточное количество разнообразных белков, которое обеспечивает существование на нашей планете не только известных ныне биологических видов (более 10 млн.), но и предусматривает возможность возникновения в ходе биологической эволюции новых видов организмов. Номенклатура пептидов
Как видно из схемы образования пептидов, полипептидная цепь имеет определенное направление, поскольку каждый из ее строительных блоков (аминокислотных звеньев, или остатков аминокислот) имеет разные концы – либо N-амино, либо С-карбоксильная группа. Аминокислотный остаток, находящийся на том конце молекулы, где имеется свободная α-аминогруппа, называется N-концевым остатком (N-конец молекулы), а остаток на противоположном конце, несущем свободную карбоксильную группу, – С-концевым (С-конец молекулы). Например, для трипептида: N – конец → (H2N) ала – гли – три (OH) ← С-конец (начало цепи) (конец цепи) Условно принято считать, что полипептидная цепь начинается с N-конца, т.е. конца, несущего α-аминогруппу. Для условной записи пептидов используют трехбуквенные сокращения сокращенные обозначения α-аминокислот (см., с.5). При изображении последовательности аминокислот в пептидной цепи начинают с N-концевого остатка. Так, в трипептиде ала – гли – три аланин несет концевую аминогруппу, а триптофан – концевую карбоксильную группу. Обратите внимание, что три – гли – ала – это уже другой трипептид! Названия пептидов образуют из тривиальных названий аминокислотных остатков в соответствии с их последовательностью, начиная с N-концевого остатка. С-концевой остаток сохраняет свое тривиальное название. Названия аминокислотных остатков производят из названий соответствующих аминокислот путем замены их окончания на окончание «ил», характерного для названий одновалентных остатков (радикалов). Например, глиц ин (остаток – глиц ил), вал ин (остаток – вал ил), фенилалан ин (остаток – фенилалан ил), аспараг ин (остаток – аспараг ил), аспарагин овая кислота (остаток – аспарагин ил) и т.д. Название трипептида ала – гли – три будет следующим: аланил – глицил – триптофан. Если аминокислотный остаток повторяется в цепи более одного раза подряд, то перед его названием используют соответствующую приставку: 2 раза – ди, 3 – три, 4 – тетра и т.д. Например, пентапептид лиз – (про)2 – тир – глн будет иметь название: лизил – дипролил – тирозил – глутамин. При наличии в пептиде в качестве С-концевого остатка глутаминовой или аспарагиновой кислоты, их называют одним словом – глутамат и аспартат соответственно. Например, название тетрапептида про – иле – асп – асп будет следующим: пролил – изолейцил – аспарагинил – аспартат.
Номенклатура ферментов В 1961 г. Международная комиссия по номенклатуре ферментов разработала научную номенклатуру. Согласно этой номенклатуре название фермента составляют из химического названия субстрата и названия той реакции, которая осуществляется ферментом. Если химическая реакция, ускоряемая ферментом, сопровождается переносом группировки атомов от субстрата к акцептору, название фермента включает также химическое название акцептора. Например, сукцинатдегидрогеназа (СДГ) (сложный фермент, простетическая группа флавиновая – ФАД), катализирующая окисление путем дегидрирования янтарной кислоты, называется: Решение упражнений Задание 1. Фермент цикла Кребса малатдегидрогеназа является НАД-зависимым ферментом, катализирующим превращение яблочной кислоты (малат) в соответствии со следующей схемой: L – малат + НАД+ → оксалоацетат + НАДН…Н+ Напишите уравнение реакции и определите к какому классу и подклассу ферментов относится малатдегидрогеназа. Решение. В предметном указателе учебника по названию фермента находим соответствующий раздел и реакцию, которую он ускоряет: COOH COOH | |
| акцептор (ox) | акцептор (red) CH2COOH CH2COOH донор (red) донор (ox) Далее даем характеристику ферменту и определяем его принадлежность к определенному классу и подклассу в соответствии с Международной классификацией ферментов. Малатдегидрогеназа является двухкомпонентным ферментом. В качестве кофермента содержит никотинамидадениндинуклеотид (НАД+), который за счет связывания со специфической белковой частью, обладает способностью восстанавливаться по ядру никотинамида (остаток витамина В5 (РР) – составная часть НАД+). В результате пиридинпротеины способны отнимать от субстратов (доноров) атом водорода, окисляя их путем дегидирования. Пиридинпротеины относятся к анаэробным дегидрогеназам, т.е. не передают снятые с субстратов атомы водорода не на кислород, а посылают их на ближайший в окислительно-восстановительной цепи другой фермент (см. раздел «Биологическое окисление»). Малатдегидрогеназа относится в первому классу (оксидоредуктазы) и к первому подклассу (окисление спиртовых групп – CH – OH), следовательно, КФ. 1.1. | Задание 2. Некоторые α-аминокислоты играют доминирующую роль в обеспечении работы мозга, так как являются предшественниками нейромедиаторов и гормонов. При отсутствии в пище такой незаменимой кислоты, как фенилаланин, у человека замедляется реакция, и он начинает «туго соображать». За способность к концентрации внимания и повышение общего тонуса отвечает дофамин (ДОФА, или 3,4-дигидроксифенилаланин), предшественником которого является аминокислота тирозин. Тирозин в свою очередь образуется из фенилаланина при каталитическом участии фенилаланин-4-гидрокслазы в соответствии со следующей схемой процесса: фенилаланин + О2 +НАДФН…Н+ → тирозин + НАДФ+ +Н2О Напишите уравнение реакции, определите класс и подкласс, к которому относится указанный фермент в соответствии с Международной классификацией ферментов. Решение. Находим в учебнике реакцию, которую катализирует указанная гидроксилаза:
│ │ CH2 CH2
OH донор акцептор донор донор донор акцептор (red) (ox) (red) (ox) (ox) (red) прямое Как видно из уравнения реакции, окисление фенилаланина осуществляется за счет прямого включения в 4-е положение бензольного ядра атома кислорода. Кроме того, в качестве парного донора, от которого поступают атомы водорода на один из атомов молекулярного кислорода, фермент использует восстановленный по ядру никотинамида кофермент НАДФН…Н+ (никотинамидадениндинуклеотидфосфат). Следовательно, указанная гидроксилаза относится к первому классу (оксидоредуктазы) и к 14 подклассу (включение одного атома водорода в два окисляемых субстрата), КФ. 1.14. Задание 3. Глутаматдекарбоксилаза катализирует превращение глутаминовой кислоты по ее α-COOH группе в соответствии со следующей схемой реакции: глутаминовая кислота → γ-аминомаслянная кислота + CO2 (ГАМК) ГАМК накапливается в мозговой ткани и действует как сигнальное вещество – нейромедиатор, тормозящий деятельность центральной нервной системы при чрезмерном психическом возбуждении. Напишите уравнение реакции, определите класс и подкласс глутаматдекарбоксилазы. Решение. Находим в учебнике реакцию, которую катализирует указанный фермент:
| (CH2)2COOH А – В А В Даем характеристику ферментам, катализирующим промежуточный обмен аминокислот путем реакций декарбоксилирования. Определяем принадлежность глутаматдекарбоксилазы к определенному классу и подклассу в соответствии с Международной классификацией ферментов. Реакции декарбоксилирования в отличие от других процессов промежуточного обмена аминокислот являются необратимыми. Они катализируются ферментами, отличающимися от декарбоксилаз α-кетокислот как по белковому компоненту, так и по природе добавочной группы. Декарбоксилазы аминокислот состоят из белковой части, обеспечивающей специфичность действия, и простетической группы, представленной пиридоксальфосфатом (производное витамина В6, см. учебник). Декарбоксилазы катализируют отщепление углекислого газа за счет разрыва в субстрате А–В связи С–С, следовательно глутаматдекарбоксилаза в соответствии с общей схемой действия (см. классификацию) относится к четвертому классу (лиазы) и первому подклассу (расщепляют связи С-С), т.е. КФ. 4.1. Задание 4. В процессе поэтапного распада пищевых белков в желудочно-кишечном тракте принимают участие различные ферменты желудочного, панкреатического и кишечного соков. Трипсин, содержащийся в панкреатическом соке, расщепляет фрагменты молекул белков после действия пепсина желудочного сока. Предположим, что после действия пепсина образовались разнообразные полипептиды (пептоны), в их числе франмент следующего полипептида: … -глу-три-ала-арг-цис-гли-гли-про-лиз-тир-гис-ала- … Приведите схему дальнейшего расщепления этого фрагмента с участием трипсина. Укажите к какому классу и подклассу относится трипсин в соответствии с Международной классификацией ферментов. Решение. По тривиальному названию фермента находим в учебнике информацию о последовательности действия ферментов желудочно-кишечного тракта в процессе переваривания белков и их избирательности (специфичности) действия после определенных аминокислот в молекуле белка. Распад белков с участием протеолитических ферментов желудочно-кишечного тракта осуществляется за счет разрыва пептидных связей с присоединением элементов воды. В соответствии с общей схемой действия (см. классификацию ферментов) указанные ферменты относятся к третьему классу (гидролазы) и четвертому подклассу (пептидазы) – гидролизуют пептидные связи (– CO – NH –), т.е. КФ. 3.4. Трипсин относится к группе протеолитических ферментов, катализирующих гидролиз пептидных связей внутри пептидной цепи. Такие ферменты получили название эндопептидаз. Эндопептидазы обладают разной субстратной специфичностью действия, определяемой природой радикалов аминокислот по соседству с разрываемой пептидной связью. Трипсин обладает более узкой субстратной специфичностью по сравнению с пепсином желудочного сока, так как разрывает пептидные связи, в образовании которых участвуют α-карбоксильные группы лизина и аргинина, т.е. основные аминокислоты (см. классификацию аминокислот). Следовательно, схему действия трипсина на фрагмент белка можно показать следующим образом: ↓ ↓ … -глу-три-ала-CONH – CH – CO – NH-цис-(гли)2-про-CONH –CH – NH-тир-гис-ала-… | | (CH2)3 (CH2)4 | | NH NH3 + | + + NH3 – C = NH2 + 2H2O → …-глу-три-ала-арг(OH) + (H2N)цис-(гли)2-про-лиз(OH) + (H2N)тир-гис-ала-... Специфичность действия трипсина нашла широкое применение в исследовательской работе при установлении первичной структуры индивидуальных белков по методу Эдмана. НУКЛЕИНОВЫЕ КИСЛОТЫ
В настоящее время нуклеиновые кислоты (НК) рассматриваются как природные вещества, которые по своей биологической значимости приравниваются к белкам. Нуклеиновые кислоты являются информационными биополимерами, так как осуществляют хранение и передачу генетической информации во всех живых организмах. Различают два типа нуклеиновых кислот: ДНК (дезоксирибонуклеиновые кислоты), которые обеспечивают сохранение генетической информации в ряду поколений в процессе репликации; РНК (рибонуклеиновые кислоты), которые обеспечивают поток генетической информации в клетке в процессе транскрипции ДНК, и трансляцию РНК (биосинтез белков). Рис. 1. Химический состав нуклеиновых кислот
Нуклеотиды являются фосфорными эфирами нуклеозидов. Нуклеозиды – это природные гликозиды азотистых оснований (пиримидиновых и пуриновых), которые связаны с пентозами через атом азота (пиримидины через N1-атом, пурины – через N9-атом (рис. 1. Б)). В зависимости от природы пентозы (углеводного остатка) различают рибонуклеозиды и дезоксирибонуклеозиды. Рис. 2. Первичная структура нуклеиновых кислот
· Полинуклеотидные цепи ДНК и РНК несут множество фосфатных групп, которые легко диссоциируют в физиологических условиях, вследствие чего молекулы НК приобретают суммарный отрицательный заряд (проявляют кислотные свойства, отсюда происхождение их названия – кислоты). В связи с кислотным характером эти соединения во многих случаях связываются в клетке с основными белками, заряженными «+», с образованием нуклеопротеиновых комплексов – рибонуклеопротеиновых (РНП) и дезоксирибонуклеопротеиновых (ДНП). · Полинуклеотидная цепь имеет определенное направление, т.е. характеризуется полярностью, поскольку все межнуклеотидные связи ориентированны вдоль цепи одинаково (3′ → 5′ фосфатные мостики). На одном конце цепи остается свободной 5′ – ОН-группа, этерифицированная фосфатом, (начало цепи); на другом – 3′ – ОН-группа, которая не связана с другим нуклеотидом (конец цепи) (рис. 1). Последовательность азотистых оснований записывается в направлении от 5′р-конца цепи к 3′(ОН) концу (аминокислотная последовательность полипептидной (белковой) цепи пишется в направлении от N-концевой к С-концевой аминокислоте, см. раздел «Пептиды» данной разработки). Например, тринуклеотиды АЦГ и ГЦА – разные соединения, точно так же, как трипептид ала-гли-три отличается от три-гли-ала. Специфические длинные последовательности азотистых оснований А, Т, Г и Ц в ДНК служат символами для кодирования генетической информации. Например, фрагмент цепи ДНК, имеющий ориентацию фосфатных мостиков 3′ → 5′, можно записать как А-Г-Ц-Ц-Т-...., или 5р А-Г-Ц-Ц-Т-..... 3(ОН), где р – остаток фосфата.
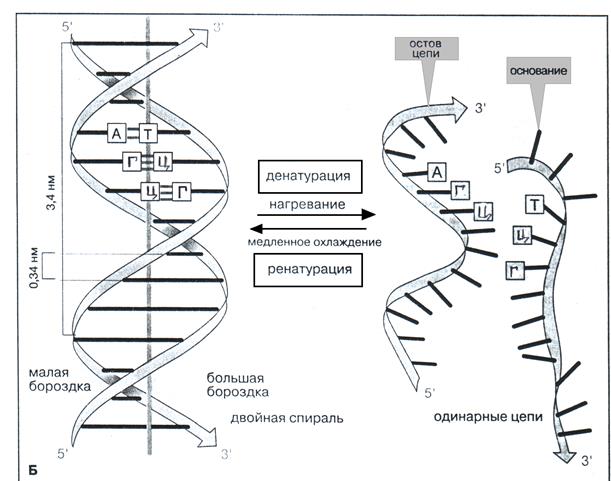
Вторичная структура ДНК Молекула ДНК в большинстве случаев состоит из двух взаимозакрученных полинуклеотидных цепей и представляет собой двойную спираль, в которой пуриновые и пиримидиновые основания направлены внутрь (рис. 3 Б). Биологическая функция молекул ДНК связана главным образом со способностью азотистых оснований образовывать специфически (комплементарно) связанные водородными связями пурин-пиримидиновые пары, принадлежащие двум цепочкам: аденин – тимин, образующие между собой две водородные связи, и гуанин – цитозин, связанные тремя водородными связями (рис.3А). Специфичность спаривания азотистых оснований обусловливает комплементарность, т.е. взаимодополняемость цепей ДНК друг к другу. Так, если в одной цепи ДНК находится сочетание азотистых оснований ТЦГА, то во второй цепи ему соответствует последовательность АГЦТ.
1. А/Т – спаривание 2. Г/Ц – спаривание
А. Спаривание оснований ДНК
Рис. 3. Вторичная структура ДНК Спаривания оснований А/Т и Г/Ц и структура в виде двойной спирали ДНК возможны только в том случае, если цепи ДНК располагаются антипараллельно, т.е. имеют противоположную направленность (полярность). Цепь, имеющую ориентацию фосфатных мостиков 3′ → 5′, т.е. полярность 5′р---...3′(ОН), называют правой и обозначают как «+» цепь. Цепь с противоположной ориентацией фосфатных мостиков 5′ → 3′, т.е. полярностью (ОН)3′----.....р5′ называют левой и обозначают как «–» цепь (рис.4). Обратите внимание, что цепи ДНК отличаются друг от друга по составу азотистых оснований, а также по их последовательности, если каждую цепь читать в направлении от 5′р-конца к 3′(ОН)-концу. Кроме того, соблюдается равенство А и Т, Г и Ц (в каждом типе ДНК содержится примерно одинаковые количества А и Т, тоже самое относится к Г и Ц).
(+) 5′рА-А-Ц-Ц-Т- Г-А-Ц-Ц-А-Ц-Т- Г-А-Т-....3′(ОН) Фрагмент (-) (ОН)3′ Т-Т- Г-Г-А-Ц-Т- Г- Г-Т- Г-А-Ц-Т-А-....р′5 ДНК Принцип антипараллельности цепей ДНК имеет глубокий биологический смысл, так как в функциональном отношении две цепи не эквивалентны. Кодирующей цепью (матричной, смысловой) является (+) цепь, которая узнается транскрибирующим ферментом (РНК-полимеразой) и считывается в процессе транскрипции ДНК. В зависимости от длины витка и расположения азотистых оснований по отношению к оси двойной спирали (угла спирали), различают более десяти разнообразных упорядоченных спиральных структур ДНК. Преобладающая в клетке конформация ДНК (так называемая В-ДНК) представлена на рис. 3Б. Длина витка (шаг спирали) составляет 3,4 нм, угол спирали - 90°. В стабилизации вторичной структуры ДНК наряду с водородными связями (Н-связи) большую роль играют гидрофобные взаимодействия между пространственно сближенными плоскостями азотистых оснований, направленные вдоль оси двойной спирали. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, из называют стэкинг-взаимодействиями (в В-форме ДНК расстояние между плоскостями азотистых оснований составляет 0,34 нм, т.к. в витке находится 10 нуклеотидных пар). Сильное стэкинг-взаимодействие всегда усиливает Н-связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные фосфатные группы которого находятся на поверхности спирали. Внутренняя область двойной спирали, где расположены основания, неполярна. При ослаблении сил стэкинг- взаимодействия молекулы воды, проникая внутрь двойной спирали, конкурируют с полярными группами оснований за образование Н-связей, что в свою очередь, приводит к нарушению Н-связей между комплементарными парами оснований и уменьшению эффективности стэкинг-взаимодействия между азотистыми основаниями, инициирует дестабилизацию двойной спирали и способствует расхождению цепей (денатурации ДНК). Ферментативное локальное расхождение цепей ДНК имеет место в генетических процессах, связанных с воспроизведением и экспрессией наследственной информации. После окончания воздействия компонентов окружающего раствора цепи ДНК, благодаря спариванию оснований, вновь образуют двойную спираль в участке их расхождения (ренатурация ДНК). Все это свидетельствует о динамичности вторичной структуры ДНК. Процессы денатурации и ренатурации ДНК играют важную роль в генной инженерии.
Рис. 4. Природа генетической информации (цепи ДНК комплементарны и антипараллельны) Вторичная структура РНК Молекулы РНК имеют одноцепочечную структуру, за исключением РНК некоторых вирусов. Из-за пространственных (стерических) препятствий, обусловленных наличием 2′– ОН-группы остатка рибозы, молекулы РНК не могут образовывать структур, подобных двойной спирали. Поэтому РНК имеют менее регулярную структуру по сравнению с ДНК, а их нуклеотидный состав не подчиняется правилу комплементарности. В большинстве молекул РНК содержание аденина (А) отличается от содержания урацила (У), а содержание гуанина (Г) отличается от содержания цитозина (Ц). Вторичная структура молекул РНК характеризуется частичной спирализацией полинуклеотидной цепи самой на себя. Биспиральные участки, называемые как «шпильки», возникают за счет образования Н-связей в тех зонах, где расположены комплементарные азотистые основания аденин – урацил (две Н-связи) и гуанин – цитозин (три Н-связи):
«шпилька»
|| || Петля» (однотяжный участок) Ц- А- Ц-Ц- У- Г- Г У | Ц А А –... 3′(ОН) В результате вторичная структура молекул РНК содержит биспиральные участки, содержащие от 3-х до 7 нуклеотидных пар, и петли, число и размеры которых определяются первичной структурой молекулы и составом окружающего раствора. Комплементарность цепей и последовательность азотистых оснований в остатках нуклеотидных звеньев составляют химическую основу важнейших функций нуклеиновых кислот: ДНК – хранение и передача наследственной информации, а РНК – непосредственное участие в биосинтезе белков. Решение упражнений Участок родительской молекулы ДНК имеет следующий нуклеотидный состав:... ТААГГАЦААГТ... .... АТТЦЦТГТТЦА... Какой нуклеотидный состав будут иметь дочерние молекулы ДНК после одного цикла репликации, если она протекает по полуконсервативному механизму?
молекула ДНК (-) 3′... АТТЦЦТГТТЦА...р′5
дочерние молекулы ДНК после одного цикла репликации
Далее необходимо дать краткую информацию о сущности процесса репликации ДНК и его основных принципах, составляющих химическую основу биологических функций ДНК; объяснить сущность полуконсервативного способа репликации и привести его экспериментальные доказательства. Для объяснения задания необходимо проработать изложенный выше теоретический материал (раздел 3), а также материал учебника «Репликация ДНК, ее механизм и способы».
· Экспрессия генетической информации
транскрипция р-РНК трансляция
генов т-РНК
Первый этап реализации наследственной информации осуществляется в процессе транскрипции генов.
Генетический код
©2015- 2026 zdamsam.ru Размещенные материалы защищены законодательством РФ.
|





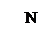
 │
H
│
H
 │ N
║ ║
│ N
║ ║

 N
│
H
N
│
H
 α
α
 α α α α
α α α α H2N–CH2–COOH
H2N–CH2–COOH H2N–CH2 –CO–NH–CH–CO –NH–CH2 – COOH и т.д.
H2N–CH2 –CO–NH–CH–CO –NH–CH2 – COOH и т.д. α
α  α
α H2N –CH–CO OH + H N –CH–COOH H2N–CH – CO – N – CH – COOH
H2N –CH–CO OH + H N –CH–COOH H2N–CH – CO – N – CH – COOH H O – C – H + НАД+ C = O + НАД Н…H +
H O – C – H + НАД+ C = O + НАД Н…H +
 H2NCHCOOH + O2 + НАДФН…Н+ → H2NCHCOOH + НАДФ+ + H2O
H2NCHCOOH + O2 + НАДФН…Н+ → H2NCHCOOH + НАДФ+ + H2O


 │
│
 H2N – CH – COO H → H2N – CH2 – CH2 – CH2 – COOH + CO2
H2N – CH – COO H → H2N – CH2 – CH2 – CH2 – COOH + CO2






 У
У 5′р Г- У- Г- Г- А- Ц- Ц У
5′р Г- У- Г- Г- А- Ц- Ц У
 Н-связи
Н-связи
 Решение. Поскольку одним из двух основных принципов строения молекулы ДНК является принцип антипараллельности цепей, то необходимо, используя условные обозначения, указать полярность цепей родительской молекулы ДНК: Дочерние цепи
Решение. Поскольку одним из двух основных принципов строения молекулы ДНК является принцип антипараллельности цепей, то необходимо, используя условные обозначения, указать полярность цепей родительской молекулы ДНК: Дочерние цепи
 (+) 5′р... ТААГГАЦААГТ...3′
(+) 5′р... ТААГГАЦААГТ...3′

 (-) 3′... АТТЦЦТГТТЦА...р′5
(-) 3′... АТТЦЦТГТТЦА...р′5
 (+) 5′р..ТААГГАЦААГТ...3′
(+) 5′р..ТААГГАЦААГТ...3′
 (-) 3′... АТТЦЦТГТТЦА....р′5
(-) 3′... АТТЦЦТГТТЦА....р′5 Под термином «экспрессия» понимают выражение генетической информации в фенотипе (фенотип – совокупность внешних признаков организма, которые формируются на основании генотипа (совокупности генов организма) и среды). Экспрессия генетической информации, содержащейся в генах (участках ДНК), реализуется с участием нуклеиновых кислот по схеме:
Под термином «экспрессия» понимают выражение генетической информации в фенотипе (фенотип – совокупность внешних признаков организма, которые формируются на основании генотипа (совокупности генов организма) и среды). Экспрессия генетической информации, содержащейся в генах (участках ДНК), реализуется с участием нуклеиновых кислот по схеме:
 ДНК м-РНК Белок
ДНК м-РНК Белок



