|
|
Принципы регуляции ферментативной активности. ⇐ ПредыдущаяСтр 3 из 3 Ряд факторов среды (температура, рН, концентрация субстратов, ингибиторов и активаторов) могут изменять скорость ферментативных реакций и в известной степени служить регуляторными факторами. Однако живые организмы не могут быстро изменять силу воздействия этих факторов и предпочитают с помощью различных регуляторных механизмов поддерживать эти факторы на определенных уровнях. Этот процесс относится к понятию гомеостаза. Гораздо чаще регуляция активности ферментов осуществляется за счет изменения количества ферментных молекул или каталитических свойств ферментов. Hразличают несколько видов регуляции: • НЕЙРОГУМОРАЛЬНАЯ регуляция (с участием центральной нервной системы, классических гормонов и гормонов местного действия) • РЕГУЛЯЦИЯ НА ГЕНЕТИЧЕСКОМ УРОВНЕ - изменение скорости биосинтеза белка. • АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ФЕРМЕНТОВ. Это регуляция, которая происходит благодаря только самим участникам реакции. Фермент не только работает, но еще и сам себя регулирует. Механизмы автономной саморегуляции очень многочисленны, но построены на двух основных принципах. 1) Механизмы, основанные на кинетических свойствах фермента, количественно характеризуемых Км и Vmax. 2) Второй принцип связан с аллостерическими свойствами фермента, то есть со способностью фермента угнетаться или активироваться под действием субстрата и/или продуктов.
Различают прямую аллостерическую регуляцию и косвенную. Под прямой аллостерической регуляцией понимают изменения в третичной структуре белковой молекулы под действием аллостерического модификатора. Под косвенной аллостерической регуляцией понимают изменение степени диссоциации белковых комплексов, образующих четвертичную структуру. Аллостерические модификаторы могут вызывать увеличение скоростей ферментативных реакций или замедление. Очень часто в качестве аллостерических модификаторов могут выступать субстраты и продукты ферментативных реакций.
НЕРАЗВЕТВЛЕННЫЕ МУЛЬТИФЕРМЕНТНЫЕ СИСТЕМЫ В большинстве случаев тот или иной биохимический процесс катализируется не одним ферментом, а целой совокупностью ферментов, каждый из которых катализирует свой этап в длинной последовательности отдельных реакций этого процесса. Совокупность ферментов, катализирующих последовательные реакции единого процесса, называется мультиферментной системой.
E1 E2 E3 E4 En А---->В----->C----->D------>....L----->M
В большинстве случаев величины Vmax у всех ферментов данной системы различны. Значит, у какого-то из ферментов Vmax меньше Vmax любого другого фермента. Такой фермент (с низкой Vmax) называется ЛИМИТИРУЮЩИМ ферментом данной мультиферментной системы. Именно этот фермент определяет скорость процесса в целом. Как правило, лимитирующими являются те ферменты, которые находятся в начале данной мультиферментной системы. Эти ферменты подвергаются воздействию регуляторных воздействий со стороны аллостерических эффекторов (в рамках автономной саморегуляции). В таких мультиферментных системах аллостерическими эффекторами бывают часто не только продукт или субстрат данной реакции, но и другие метаболиты - в особенности конечные продукты (М). Если конечный продукт оказывает активирующее действие, то этот механизм называется положительной обратной связью и процесс протекает взрывообразно - до полного исчерпания субстрата в системе. Если конечный продукт - ингибитор лимитирующего фермента - то говорят об отрицательной обратной связи, и при увеличении концентрации конечного продукта скорость процесса будет замедляться. Если лимитирующий фермент является одновременно и регуляторным, то его называют КЛЮЧЕВЫМ ферментом данной мультиферментной цепи.
РАЗВЕТВЛЕННЫЕ МУЛЬТИФЕРМЕНТНЫЕ СИСТЕМЫ. Часто мультиферментные цепи являются разветвленными. В таких системах ключевые ферменты обычно предшествуют участкам ветвления, а сразу за разветвлением находятся ПУНКТЫ ВТОРИЧНОГО КОНТРОЛЯ, то есть такие ключевые ферменты, которые регулируют скорость реакций в своей ветви:
В этом примере Е1 - ключевой фермент, а Е4 и Е7 - пункты вторичного контроля.
Активирование Чаще всего в момент образования, в силу ряда причин, фермент оказывается малоактивным и приведение его в активное состояние осуществляется главным образом следующими путями: 1. Высвобождение активного центра путем отщепления олигопептида закрывающего его. Например, соляная кислота отщепляет олигопептид закрывающий активный центр пепсиногена переводя его в пепсин. 2. Активация ионами - ионы металлов участвуют в образовании фермент-субстратного комплекса. На первой стадии образуется комплекс Ме-субстрат, на второй – субстрат фермент. 3. Аллостерическая активация – действие специальных веществ (модуляторы или эффекторы) на аллостерический центр вызывают конформационные изменения в белковой структуре фермента, тем самым активируя активный центр. Как правило аллостерический и активный центр отдалены друг от друга.
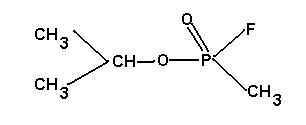
Ингибирование. Вещества, которые блокируют каталитическую активность фермента называются ингибиторами. При взаимодействии ингибитора с ферментом образуется комплекс, прочность которого определяет тип ингибирования: обратимое или необратимое. Необратимое ингибирование: прочный комплекс, при котором фермент навсегда теряет каталитическую активность. Необратимое ингибирование отдельных ферментов это смерть живого организма. Типичным примером необратимомого ингибитора может служить фосфоорганическое соединение диизопропилфторфосфат (ДФФ), которое ингибирует фермент ацетилхолинэстеразу.
Этот фермент играет важную роль при передаче нервных импульсов, расщепляя нейромедиатор ацетилхолин. Ингибитор, обладая высокой реакционной способностью, присоединяется к остатку серина в активном центре ацетилхолинэстеразе образуя неактивное производное. Фермент перестает «работать» и организм погибает от нервного перевозбуждения. Фософорганические соединения очень токсично для нервной системы и получили название «нервные яды». Помимо ДФФ к этому классу можно отнести: Тетроэтилпирофосфат:
Зарин
Табун.
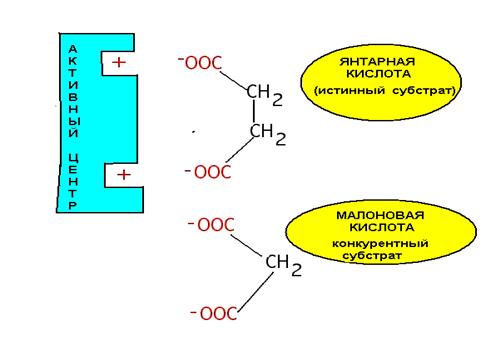
Обратимое ингибирование отличается от необратимого тем, что каталитическую активность фермента можно возобновить. Обратимое ингибирование делится на две группы: конкурентное и неконкурентное. Конкурентное - ингибитор и субстрат имеют похожее строение. В этом случае ингибитор, присоединяясь к активному центру, не может участвовать в химической реакции. Повышение концентрации истинного субстрата снимает ингибитор с активного центра. Примером может служить малоновая кислота, которая угнетает работу сукцинатдегидрогеназы.
Практическим примером применения конкурентного ингибирования является использование сульфаниламидных препаратов в медицине. Строение сульфаниламида напоминает парааминбензойную кислоту, которую используют бактерии для синтеза необходимой для их жизнедеятельности фолиевой кислоты. Благодаря этому структурному сходству сульфаниламид блокирует активный центр фермента синтезирующего фолиевую кислоту, что приводит к замедлению роста бактерий.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами ферментов. Ингибитор соединяется с ферментом не в активном центре, и данное присоединение ведет к конформационным изменениям всей белковой структуры. Примером может служить действие ионов тяжелых металлов. Но существуют так называемые «суицидные субстраты», которые по своему строению напоминают истинный субстрат, но имеют одну или несколько реакционных группировок, которые блокируют активный центр. В этом случае неконкурентное ингибирование переходит в необратимое.
Коферменты.
Небелковая часть фермента – кофермент. Многие ферментативные реакции протекают с переносом электронов или различных функциональных групп. В таких реакциях всегда принимают участие коферменты, выполняющие роль промежуточных переносчиков электронов или функциональных групп. Ферменты обычно высокоспецифичны к своим субстратам, коферменты же взаимодействуют с разными ферментами одного класса обладающими различной субстратной специфичностью. По способам взаимодействия фермента с коферментом различают: Растворимый кофермент или кофактор, нековалентно связанный с глобулой апофермента; Простетическую группу – ковалентно связанную с апоферментом. Большинство коферментов – водорастворимые витамины. Впервые в чистом виде витамины были получены в 1912 году Казимиром Функом (витамин В1). В 1922 году Зелинским выдвинул гипотезу о биологическом действии витаминов, которая затем подтвердилась. Согласно этой гипотезе витамины участвуют и регулируют биологические процессы опосредовано через ферментные системы, в которые они ходят в качестве кофакторов или простетических групп.
Кофакторы оксидоредуктаз.
Все оксидоредуктазы нуждаются в коферментах, которые представлены в виде кофактора или простетической группы. Основная функция перенос электронов и протонов водорода (е + Н+) – электрохимический эквивалент.
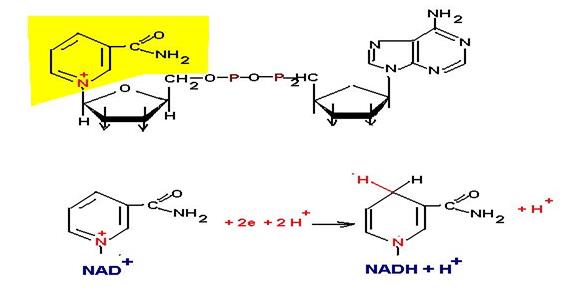
Производные никотинамида. Чаще всего представлен в виде никотинамиддинуклеотида NAD и никотинамиддинуклеотидфосфат NADF. Данный кофактор всегда присутствует в растворенной форме и переносит электрохимический эквивалент в дыхательную цепь. Причем NAD чаще всего участвует в процесса распада, а NADF в процессах биосинтеза.
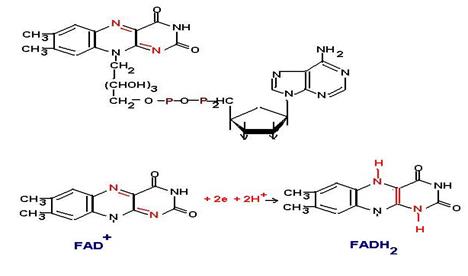
Флавиновые ферменты. Витамин В2. Являются производными флавина. Это простетическая группа прочно связана с глобулой апофермента. Представлена двумя формами флавинмононуклеотид FMN и флавинадениндинуклеотид FAD
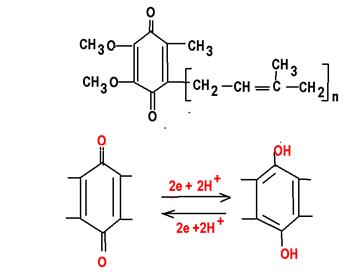
Убихинон (кофермент Q), структурный компонент дыхательной цепи
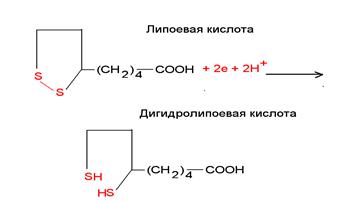
Липоевая кислота – участвует в окислительном декарбоксилировании
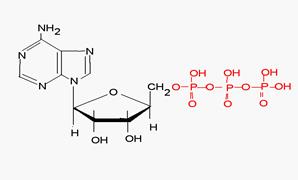
Коферменты переноса групп Нуклеозидфосфаты (АТФ, АДФ, ГТФ) переносят остатки фосфорной кислоты
Тиаминдифосфат (ТДФ). Витамин В1. Активирует альдегиды и кетоны и переносит их на другие молекулы. Участвует в работе пируватдегидрогеназной мультиферментной системы.
Пантотеновая кислота – структурный компонент кофермента А (коэнзима А) перенос ацетильных и ацильных остатков
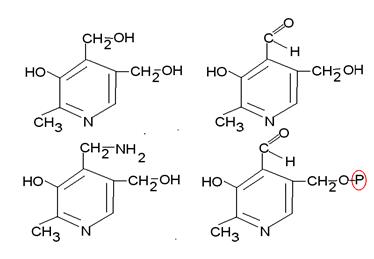
Пиридоксин. Витамин В6. Существует в нескольких формах: пиридоксаль, пиридоксинамин и пиридоксальфосфат. Участвует в реакциях переаминирования аминокислот в кетокарбоновые. Перенос аминогруппы
Биотин (витамин Н), участвует в реакциях с переносом карбоксильных групп (карбоксилирования и декарбоксилирования).
  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|