|
|
Уравнение скорости ферментативной реакции.Стр 1 из 3Следующая ⇒ ФЕРМЕНТЫ 1. Определение понятия фермент. Биохимическая природа ферментов. Функции ферментов в живом организме. Специфические черты биологического катализа. 2. Классификация ферментов. 3. Уравнение скорости ферментативной реакции. Константы ферментативной реакции Км и Vмах. Единицы ферментативной активности. 4. Активный центр. Факторы ферментативного катализа. 5. Влияние условий среды на скорость ферментативной реакции 6. Принципы регуляции ферментативной активности.. 7. Активирование и ингибирование ферментов 8. Коферменты. Определение понятия фермент. Биохимическая природа ферментов. Функции ферментов в живом организме. Специфические черты биологического катализа. Ферментами называют специализированную группу белков выполняющих в живом организме функцию кинетического контроля (контроль скорости) химических процессов. По определению Джона Бернала отличительным свойством живых организмов является их способность кинетически контролировать стремление живых систем к состоянию термодинамического равновесия. Эта функция выражается в способности ферментов к катализу, поскольку в отсутствие фермента скорости большинства химических реакций в организме практически равны нулю. Катализаторами называют химические соединения, в присутствии которых увеличивается скорость химической реакции. Катализаторы вступают во взаимодействие с реагентами, изменяя механизм химической реакции, и к моменту образования продуктов полностью восстанавливают свою структуру и химические свойства. Под механизмом химической реакции понимают совокупность промежуточных стадий при переходе от исходных веществ к конечным продуктам реакции. Для каждой стадии характерны свои промежуточные продукты (интермедиаты). Способностью к биологическому катализу обладают главным образом белки – ферменты, в меньшей степени нуклеиновые кислоты, коферменты и ряд ионов металлов. Фермент ускоряет химическую реакцию путем изменения механизма реакции. Результатом этого процесса является снижение энергии активации реакции.
Белковый катализ имеет ряд специфических свойств, связанных с химической природой фермента. 1. Высокая эффективность. Под эффективностью понимают способность катализатора поддерживать высокие скорости реакции при нормальной температуре и давлении. 2. Высокая избирательность (специфичность). Под избирательностью понимают специфичность катализатора к реагентам и расщепляемым связям. 3. Способность к регуляции каталитических свойств. Способность к регуляции выражается в способности фермента изменять скорость катализируемой реакции под воздействием факторов внешней среды, неспецифических и специфических (регуляторных молекул или при взаимодействии с другими белковыми системами).
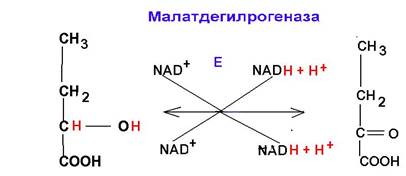
Классификация ферментов. Значительный рост числа открываемых ферментов повлек за собой трудности в их классификации. Многие исследователи присваивали вновь открытым ферментам названия, которые не отражали ни природу субстрата, ни тип ферментативной реакции (каталаза, енолаза, промежуточный фермент, рН-фермент и др.) В 1861 году комиссия по ферментам Международного биохимического союза приняла принципы систематизации и номенклатуры ферментов. В основе классификации ферментов лежит механизм катализируемой химической реакции, выраженный химическим уравнением. Первыйобщий принцип. Название фермента, включая названия с суффиксом – аза, используются для обозначения индивидуальных ферментов. Для обозначения функциональных комплексов ферментов используется название система (например, сукцинатоксидазная система). Второй общий принцип. За основу номенклатуры принимается суммарная реакция, выраженная формальным уравнением. Следствием этого принципа является то, что определенное название обозначает не индивидуальный белок, а группу белков с одинаковым каталитическим действием. Для дифференциации названий такой группы применяют дополнительные виды классификаций, основанные на дополнительных признаках. Третий общий принцип. Ферменты подразделяются на группы в соответствии с типом катализируемой реакции, и этот последний в сочетании с названием субстратов служит основой для построения названий отдельных ферментов, включая кодовую нумерацию (шифры) ферментов. Таким образом, для обозначения ферментов используют систематические названия, рабочие (тривиальные) названия, шифры ферментов. Схема классификации ферментов. Все ферменты делятся на шесть классов: 1. Оксидоредуктазы. Ферменты катализирующие окислительно-восстановительные реакции.
2. Трансферазы Ферменты, катализирующие реакции переноса групп (реакции замещения)
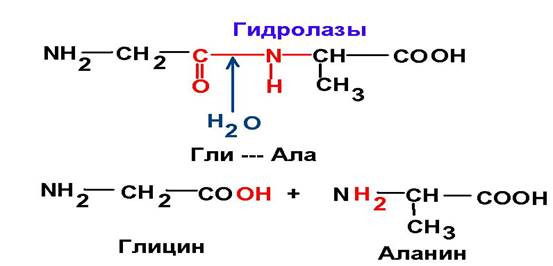
3. Гидролазы Ферменты, катализирующие реакции гидролиза.
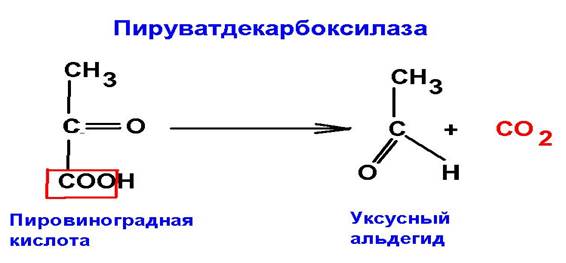
4. Лиазы. Ферменты, катализирующие реакции присоединения по двойной связи или отщепления с образованием двойной связи.
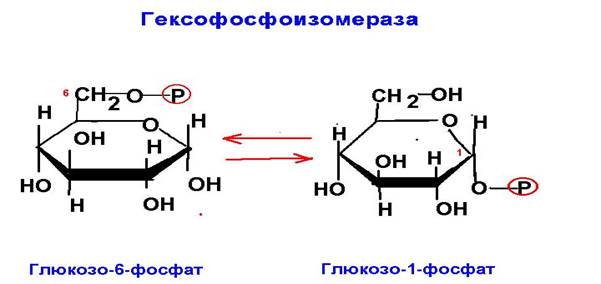
5. Изомеразы Ферменты, катализирующие внутримолекулярный перенос химических групп.
6. Лигазы (синтетазы) Ферменты, катализирующие соединение друг с другом двух молекул, сопряженное с распадом молекулы АТФ или аналогичного трифосфата.
Внутри каждого класса ферменты подразделяются на подклассы в зависимости от типа субстрата, с которым они взаимодействуют. Эти подклассы делятся на подподклассы, детализирующие природу катализирующей реакции. В соответствии с рассмотренной системой классификации каждому ферменту присваивается номер по четырехзначному десятичному коду КФ 1.1.1.1. Таким образом систематизировано более 2000 ферментов. Но в биохимической практике также остаются тривиальные названия.
Влияние температуры Зависимость скорости ферментативной реакции от температуры имеет вид оптимальной кривой с максимумом, называемым температурным оптимумом ферментативной реакции. Для химических каталитических систем аналогичная зависимость представлена монотонно возрастающей кривой. Параметром оценки температурной зависимости является величина Q10, обозначающая во сколько раз возрастает скорость реакции при увеличении температуры на десять градусов. Обычно значения параметра колеблются в пределах от двух до трех. Появление оптимума на кривой зависимости скорости ферментативной реакции от температуры связано с наложением двух процессов увеличением скорости при увеличении температуры и снижением активности фермента в результате температурной денатурации фермента. Таким образом, температурный оптимум не есть величина постоянная для данного фермента. Она зависит от времени инкубации, а также влияния компонентов реакционной среды (в первую очередь концентраций субстратов и продуктов) на скорость денатурации фермента. Поэтому для измерения активности фермента рекомендуется температура 25 С0, при которой большинство ферментов денатурирует незначительно.
Влияние рН Зависимость активности фермента от величины рН имеет также характер кривой с оптимумом, однако, причины такой закономерности иные, чем в случае зависимости от температуры. Характер влияния рН на скорость ферментативной реакции связан со степенью протонирования полярных групп активного центра фермента, участвующих в катализе.
Активирование Чаще всего в момент образования, в силу ряда причин, фермент оказывается малоактивным и приведение его в активное состояние осуществляется главным образом следующими путями: 1. Высвобождение активного центра путем отщепления олигопептида закрывающего его. Например, соляная кислота отщепляет олигопептид закрывающий активный центр пепсиногена переводя его в пепсин. 2. Активация ионами - ионы металлов участвуют в образовании фермент-субстратного комплекса. На первой стадии образуется комплекс Ме-субстрат, на второй – субстрат фермент. 3. Аллостерическая активация – действие специальных веществ (модуляторы или эффекторы) на аллостерический центр вызывают конформационные изменения в белковой структуре фермента, тем самым активируя активный центр. Как правило аллостерический и активный центр отдалены друг от друга.
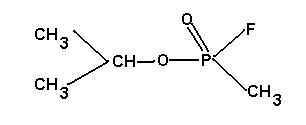
Ингибирование. Вещества, которые блокируют каталитическую активность фермента называются ингибиторами. При взаимодействии ингибитора с ферментом образуется комплекс, прочность которого определяет тип ингибирования: обратимое или необратимое. Необратимое ингибирование: прочный комплекс, при котором фермент навсегда теряет каталитическую активность. Необратимое ингибирование отдельных ферментов это смерть живого организма. Типичным примером необратимомого ингибитора может служить фосфоорганическое соединение диизопропилфторфосфат (ДФФ), которое ингибирует фермент ацетилхолинэстеразу.
Этот фермент играет важную роль при передаче нервных импульсов, расщепляя нейромедиатор ацетилхолин. Ингибитор, обладая высокой реакционной способностью, присоединяется к остатку серина в активном центре ацетилхолинэстеразе образуя неактивное производное. Фермент перестает «работать» и организм погибает от нервного перевозбуждения. Фософорганические соединения очень токсично для нервной системы и получили название «нервные яды». Помимо ДФФ к этому классу можно отнести: Тетроэтилпирофосфат:
Зарин
Табун.
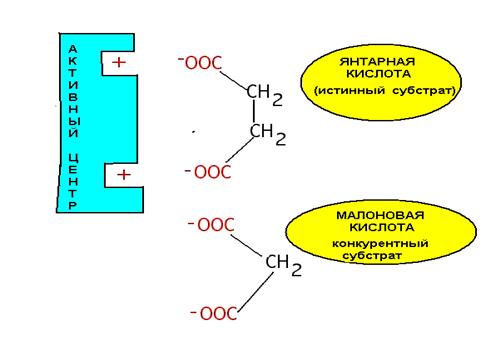
Обратимое ингибирование отличается от необратимого тем, что каталитическую активность фермента можно возобновить. Обратимое ингибирование делится на две группы: конкурентное и неконкурентное. Конкурентное - ингибитор и субстрат имеют похожее строение. В этом случае ингибитор, присоединяясь к активному центру, не может участвовать в химической реакции. Повышение концентрации истинного субстрата снимает ингибитор с активного центра. Примером может служить малоновая кислота, которая угнетает работу сукцинатдегидрогеназы.
Практическим примером применения конкурентного ингибирования является использование сульфаниламидных препаратов в медицине. Строение сульфаниламида напоминает парааминбензойную кислоту, которую используют бактерии для синтеза необходимой для их жизнедеятельности фолиевой кислоты. Благодаря этому структурному сходству сульфаниламид блокирует активный центр фермента синтезирующего фолиевую кислоту, что приводит к замедлению роста бактерий.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами ферментов. Ингибитор соединяется с ферментом не в активном центре, и данное присоединение ведет к конформационным изменениям всей белковой структуры. Примером может служить действие ионов тяжелых металлов. Но существуют так называемые «суицидные субстраты», которые по своему строению напоминают истинный субстрат, но имеют одну или несколько реакционных группировок, которые блокируют активный центр. В этом случае неконкурентное ингибирование переходит в необратимое.
Коферменты.
Небелковая часть фермента – кофермент. Многие ферментативные реакции протекают с переносом электронов или различных функциональных групп. В таких реакциях всегда принимают участие коферменты, выполняющие роль промежуточных переносчиков электронов или функциональных групп. Ферменты обычно высокоспецифичны к своим субстратам, коферменты же взаимодействуют с разными ферментами одного класса обладающими различной субстратной специфичностью. По способам взаимодействия фермента с коферментом различают: Растворимый кофермент или кофактор, нековалентно связанный с глобулой апофермента; Простетическую группу – ковалентно связанную с апоферментом. Большинство коферментов – водорастворимые витамины. Впервые в чистом виде витамины были получены в 1912 году Казимиром Функом (витамин В1). В 1922 году Зелинским выдвинул гипотезу о биологическом действии витаминов, которая затем подтвердилась. Согласно этой гипотезе витамины участвуют и регулируют биологические процессы опосредовано через ферментные системы, в которые они ходят в качестве кофакторов или простетических групп.
Кофакторы оксидоредуктаз.
Все оксидоредуктазы нуждаются в коферментах, которые представлены в виде кофактора или простетической группы. Основная функция перенос электронов и протонов водорода (е + Н+) – электрохимический эквивалент.
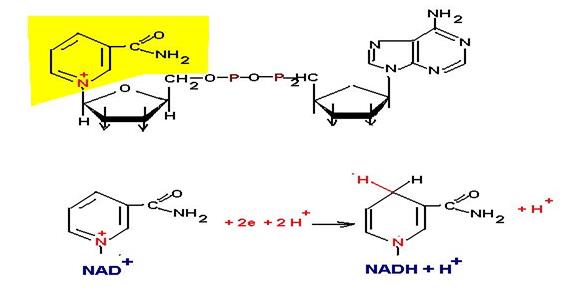
Производные никотинамида. Чаще всего представлен в виде никотинамиддинуклеотида NAD и никотинамиддинуклеотидфосфат NADF. Данный кофактор всегда присутствует в растворенной форме и переносит электрохимический эквивалент в дыхательную цепь. Причем NAD чаще всего участвует в процесса распада, а NADF в процессах биосинтеза.
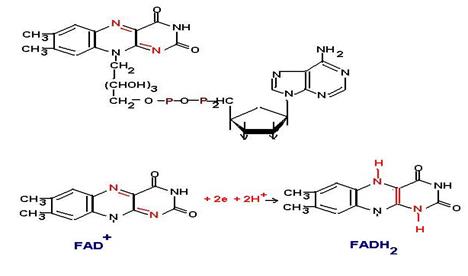
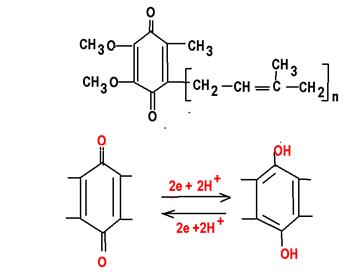
Флавиновые ферменты. Витамин В2. Являются производными флавина. Это простетическая группа прочно связана с глобулой апофермента. Представлена двумя формами флавинмононуклеотид FMN и флавинадениндинуклеотид FAD
Убихинон (кофермент Q), структурный компонент дыхательной цепи
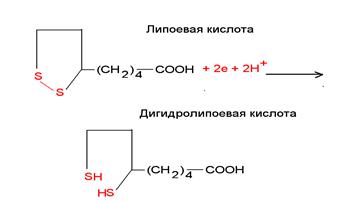
Липоевая кислота – участвует в окислительном декарбоксилировании
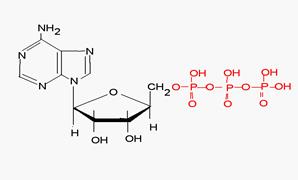
Коферменты переноса групп Нуклеозидфосфаты (АТФ, АДФ, ГТФ) переносят остатки фосфорной кислоты
Тиаминдифосфат (ТДФ). Витамин В1. Активирует альдегиды и кетоны и переносит их на другие молекулы. Участвует в работе пируватдегидрогеназной мультиферментной системы.
Пантотеновая кислота – структурный компонент кофермента А (коэнзима А) перенос ацетильных и ацильных остатков
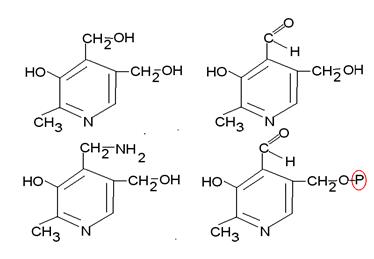
Пиридоксин. Витамин В6. Существует в нескольких формах: пиридоксаль, пиридоксинамин и пиридоксальфосфат. Участвует в реакциях переаминирования аминокислот в кетокарбоновые. Перенос аминогруппы
Биотин (витамин Н), участвует в реакциях с переносом карбоксильных групп (карбоксилирования и декарбоксилирования).
ФЕРМЕНТЫ 1. Определение понятия фермент. Биохимическая природа ферментов. Функции ферментов в живом организме. Специфические черты биологического катализа. 2. Классификация ферментов. 3. Уравнение скорости ферментативной реакции. Константы ферментативной реакции Км и Vмах. Единицы ферментативной активности. 4. Активный центр. Факторы ферментативного катализа. 5. Влияние условий среды на скорость ферментативной реакции 6. Принципы регуляции ферментативной активности.. 7. Активирование и ингибирование ферментов 8. Коферменты. Определение понятия фермент. Биохимическая природа ферментов. Функции ферментов в живом организме. Специфические черты биологического катализа. Ферментами называют специализированную группу белков выполняющих в живом организме функцию кинетического контроля (контроль скорости) химических процессов. По определению Джона Бернала отличительным свойством живых организмов является их способность кинетически контролировать стремление живых систем к состоянию термодинамического равновесия. Эта функция выражается в способности ферментов к катализу, поскольку в отсутствие фермента скорости большинства химических реакций в организме практически равны нулю. Катализаторами называют химические соединения, в присутствии которых увеличивается скорость химической реакции. Катализаторы вступают во взаимодействие с реагентами, изменяя механизм химической реакции, и к моменту образования продуктов полностью восстанавливают свою структуру и химические свойства. Под механизмом химической реакции понимают совокупность промежуточных стадий при переходе от исходных веществ к конечным продуктам реакции. Для каждой стадии характерны свои промежуточные продукты (интермедиаты). Способностью к биологическому катализу обладают главным образом белки – ферменты, в меньшей степени нуклеиновые кислоты, коферменты и ряд ионов металлов. Фермент ускоряет химическую реакцию путем изменения механизма реакции. Результатом этого процесса является снижение энергии активации реакции.
Белковый катализ имеет ряд специфических свойств, связанных с химической природой фермента. 1. Высокая эффективность. Под эффективностью понимают способность катализатора поддерживать высокие скорости реакции при нормальной температуре и давлении. 2. Высокая избирательность (специфичность). Под избирательностью понимают специфичность катализатора к реагентам и расщепляемым связям. 3. Способность к регуляции каталитических свойств. Способность к регуляции выражается в способности фермента изменять скорость катализируемой реакции под воздействием факторов внешней среды, неспецифических и специфических (регуляторных молекул или при взаимодействии с другими белковыми системами).
Классификация ферментов. Значительный рост числа открываемых ферментов повлек за собой трудности в их классификации. Многие исследователи присваивали вновь открытым ферментам названия, которые не отражали ни природу субстрата, ни тип ферментативной реакции (каталаза, енолаза, промежуточный фермент, рН-фермент и др.) В 1861 году комиссия по ферментам Международного биохимического союза приняла принципы систематизации и номенклатуры ферментов. В основе классификации ферментов лежит механизм катализируемой химической реакции, выраженный химическим уравнением. Первыйобщий принцип. Название фермента, включая названия с суффиксом – аза, используются для обозначения индивидуальных ферментов. Для обозначения функциональных комплексов ферментов используется название система (например, сукцинатоксидазная система). Второй общий принцип. За основу номенклатуры принимается суммарная реакция, выраженная формальным уравнением. Следствием этого принципа является то, что определенное название обозначает не индивидуальный белок, а группу белков с одинаковым каталитическим действием. Для дифференциации названий такой группы применяют дополнительные виды классификаций, основанные на дополнительных признаках. Третий общий принцип. Ферменты подразделяются на группы в соответствии с типом катализируемой реакции, и этот последний в сочетании с названием субстратов служит основой для построения названий отдельных ферментов, включая кодовую нумерацию (шифры) ферментов. Таким образом, для обозначения ферментов используют систематические названия, рабочие (тривиальные) названия, шифры ферментов. Схема классификации ферментов. Все ферменты делятся на шесть классов: 1. Оксидоредуктазы. Ферменты катализирующие окислительно-восстановительные реакции.
2. Трансферазы Ферменты, катализирующие реакции переноса групп (реакции замещения)
3. Гидролазы Ферменты, катализирующие реакции гидролиза.
4. Лиазы. Ферменты, катализирующие реакции присоединения по двойной связи или отщепления с образованием двойной связи.
5. Изомеразы Ферменты, катализирующие внутримолекулярный перенос химических групп.
6. Лигазы (синтетазы) Ферменты, катализирующие соединение друг с другом двух молекул, сопряженное с распадом молекулы АТФ или аналогичного трифосфата.
Внутри каждого класса ферменты подразделяются на подклассы в зависимости от типа субстрата, с которым они взаимодействуют. Эти подклассы делятся на подподклассы, детализирующие природу катализирующей реакции. В соответствии с рассмотренной системой классификации каждому ферменту присваивается номер по четырехзначному десятичному коду КФ 1.1.1.1. Таким образом систематизировано более 2000 ферментов. Но в биохимической практике также остаются тривиальные названия.
Уравнение скорости ферментативной реакции.   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|