
|
|
Принципы координации в деятельности ЦНС.
В условиях физиологической нормы работа всех органов и систем тела является согласованной: на воздействия из внешней и внутренней среды организм реагирует как единое целое. Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации. Явления координации играют важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров. За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т.д.). Установлен ряд общих закономерностей - принципов координации: принцип конвергенции; принцип иррадиации возбуждения; принцип реципрокности; принцип последовательной смены возбуждения торможением и торможения возбуждением; феномен «отдачи»; цепные и ритмические рефлексы; принцип общего конечного пути; принцип обратной связи; принцип доминанты. Разберем некоторые из них. Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, при ходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эфферентным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эфферентных, поэтому афферентные нейроны образуют на телах и дендритах эфферентных и вставочных нейронов многочисленные синапсы. Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры. Принцип реципрокности (сопряженности) в работе нервных центров. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов - сгибателей и разгибателей конечностей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы. Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное), но может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д. Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр. Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы. Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эфферентных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям. Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эфферентные нейроны). Общими путями могут быть не только конечные эфферентные нейроны, но и вставочные. Импульсы, сходящиеся в общем пути, «конкурируют» друг с другом за использование этого пути. Так достигается упорядочение рефлекторного ответа, соподчинение рефлексов и затормаживание менее существенных. Вместе с тем организм получает возможность реагировать на различные раздражения из внешней и внутренней среды при помощи сравнительно небольшого количества исполнительных органов. Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы несущие информацию о характеристиках этого мышечного сокращения вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов), получили название вторичных афферентных импульсов, или обратной связи. Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные - их угнетению. За счет положительных и отрицательных обратных связей осуществляется, например, регуляция относительного постоянства величины артериального давления. Принцип доминанты. Принцип доминанты сформулирован А. Л. Ухтомским, Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта - это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения. Доминантный очаг возбуждения характеризуется следующими основными свойствами: · повышенной возбудимостью; · стойкостью возбуждения; · способностью к суммированию возбуждения; · инерцией, доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения. Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется. Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку. Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека. Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения. Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
Спинной мозг.
Спинной мозг лежит в позвоночном канале и представляет собой тяж длиной 41 - 45 см (у взрослого), несколько сплющенный спереди назад. Вверху он непосредственно переходит в головной мозг, а внизу заканчивается заострением - мозговым конусом - на уровне II поясничного позвонка. Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде задних корешков. Морфологической границы между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным. Каждый сегмент через свои корешки иннервирует три метамера тела собственный, половину вышележащего и половину нижележащего, а каждый метамер тела получает иннервацию от трех сегментов спинного мозга. Такое устройство гарантирует осуществление функций спинного мозга при возможных его перерывах и других поражениях. Распределение функций входящих и выходящих волокон спинного мозга подчиняется определенному закону: все чувствительные (афферентные) волокна входят в спинной мозг через его задние корешки, а двигательные и вегетативные (эфферентные) выходят через передние корешки. Этот закон известен как закон Белла-Мажанди. В задних корешках волокон гораздо больше, чем в передних (их соотношение у человека примерно 5:1) т. е. при большом разнообразии поступающей информации организм использует незначительное количество исполнительных приборов. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям. Задние корешки образованы волокнами одного из отростков афферентных нейронов, тела которых расположены вне центральной нервной системы – в межпозвоночных ганглиях, а волокна другого отростка связаны с рецептором. Общее число афферентных волокон у человека достигает примерно 1 млн. Они различаются по диаметру. Наиболее толстые идут от рецепторов мышц и сухожилий, средние по толщине—от тактильных рецепторов кожи, от части мышечных рецепторов и от рецепторов внутренних органов (мочевого пузыря, желудка, кишечника и др.), наиболее тонкие миелинизированные и немиелинизированные волокна—от болевых рецепторов и терморецепторов. Одна часть афферентных волокон заканчивается на нейронах спинного мозга, другая часть направляется к нейронам продолговатого мозга, образуя спинно-бульбарный путь. Передние корешки состоят из отростков мотонейронов передних рогов спинного мозга и нейронов боковых рогов. Волокна первых направляются к скелетной мускулатуре, а волокна вторых переключаются в вегетативных ганглиях на другие нейроны и иннервируют внутренние органы. Спинной мозг имеет два утолщения: шейное и поясничное, соответствующие местам выхода из него нервов, идущих к верхней и нижней конечностям. Передней срединной щелью и задней срединной бороздкой спинной мозг делится на две симметричные половины, каждая в свою очередь имеет по две слабовыраженные продольные борозды, из которых выходят передние и задние корешки - спинномозговые нервы. Эти борозды разделяют каждую половину на три продольных тяжа - канатика: передний, боковой и задний. В поясничном отделе корешки идут параллельно концевой нити и образуют пучок, носящий название конского хвоста.
Передний рог имеет округлую или четырехугольную форму и содержит клетки, дающие начало передним (двигательным) корешкам спинного мозга. Задний рог уже и длиннее и включает клетки, к которым подходят чувствительные волокна задних корешков. Боковой рог образует небольшой треугольной формы выступ, состоящий из клеток, относящихся к вегетативной части нервной системы. Белое вещество спинного мозга составляет передний, боковой и задний канатики и образовано преимущественно продольно идущими нервными волокнами, объединенными в пучки - проводящие пути. Среди них выделяют три основных вида: · волокна, соединяющие участки спинного мозга на различных уровнях; · двигательные (нисходящие) волокна, идущие из головного мозга в спинной на соединение с клетками, дающими начало передним двигательным корешкам; · чувствительные (восходящие) волокна, которые частично являются продолжением волокон задних корешков, частично отростками клеток спинного мозга и восходят кверху к головному мозгу. От спинного мозга, образуясь из передних и задних корешков, отходит 31 пара смешанных спинномозговых нервов: 8 пар шейных, 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых. Участок спинного мозга, соответствующий отхождению пары спинномозговых нерввов, называют сегментом спинного мозга. В спинном мозге выделяют 31 сегмент. Спинной мозг выполняет две функции: рефлекторную и проводниковую. Рефлекторные центры спинного мозга. Объем функций, осуществляемых спинным мозгом, чрезвычайно велик. В шейном отделе спинного мозга находятся центр диафрагмального нерва и центр сужения зрачка, в шейном и грудном отделах - центры мышц верхних конечностей, мышц груди, спины и живота, в поясничном отделе - центры мышц нижних конечностей, в крестцовом отделе - центры мочеиспускания, дефекации и половой деятельности, в боковых рогах грудного и поясничного отделов спинного мозга - центры потоотделения и спинальные сосудодвигательные центры. В естественных условиях эти рефлексы всегда испытывают влияние высших отделов головного мозга. Степень проявления рефлексов зависит от того, сохраняются ли связи структур спинного мозга со структурами головного мозга. После децеребрации (удаления головного мозга) или спинализации (отделения спинного мозга от головного посредством перерезки) исчезают многие сложные формы активности, создаваемые спинным мозгом. При этом определенное значение принадлежит уровню организации подопытного животного. Например, спинальная лягушка, может сидеть и вырываться, когда ее схватывают, собака же сама не может ни стоять, ни ходить. Причина этого в разобщении спинного мозга и вышележащих структур нарушает. При этом, в частности, исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные движения, пропадают тонические разряды симпатических нейронов, поддерживающих сосудистый тонус и соответственно артериальное давление. В зависимости от числа нейронов, участвующих в проведении возбуждения, рефлекторные дуги спинного мозга делятся на моносинаптические и полисинаптические. За исключением моносинаптических рефлексов растяжения рефлекторные дуги всех остальных спиномозговых рефлексов являются полисинаптическими.
Защитные рефлексы запускаются, как правило, с рецепторов кожи, хотя характер ответной реакции сильно зависит от силы и вида раздражителя. Чаще всего конечная реакция выглядит в виде усиления сокращения мышц сгибателей. Рефлексы растяжения проявляются укорочением мышцы в ответ на ее растяжение. Основными рецепторами в этом случае служат нервно-мышечные веретена, афферентным звеном чувствительные волокна соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны с мышцей. Рефлексы наиболее выражены в мышцах-разгибателях. Для того. чтобы организм мог противостоять силе земного притяжения, эти мышцы должны находиться в состоянии тонического напряжения. физиологическое значение этих рефлексов состоит в том, что они участвуют в сохранении статики и положения тела, регулируя степень сокращения мышцы в соответствии с падающими на нее раздражениями. Рефлексы мышц-антагонистов лежат в основе локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей одновременно происходит торможение мотонейронов мыщц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. В целом это создает правильное чередование противоположных по функциональному значению мышечных сокращений. Механизм, обусловливающий такое чередование активности различных двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате спинного мозга. Вместе с тем для его активации необходимо поступление тонического нисходящего сигнала из двигательных центров головного мозга. Висцеромоторные рефлексы возникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины. Возникновение этих рефлексов связано с существованием конвергенции висцеральных и соматических аффферентных волокон к одним и тем же интернейронам спинного мозга. Вегетативные рефлексы заключаются, во-первых, в появлении полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на возбуждение симпатических и соматических чувствительных клеток, во-вторых, в возникновении рефлекторных реакций парасимпатических нейронов в ответ на раздражение чувствительных путей. Наряду с выполнением собственных рефлекторных реакций нейронные структуры спинного мозга служат аппаратом для реализации большого числа сложных процессов, осуществляемых различными отделами головного мозга. Проводниковая функция спинного мозга. Через спинной мозг проходят восходящие и нисходящие нервные пути. Восходящие нервные пути передают информацию от тактильных, болевых, температурных рецепторов кожи и от проприорецепторов мышц через нейроны спинного мозга в вышележащие отделы центральной нервной системы, к мозжечку и горе головного мозга. Нисходящие нервные пути (пирамидный и экстрапирамидный) связывают кору головного мозга, подкорковые ядра и образования ствола мозга с мотонейронами спинного мозга. Они обеспечивают влияние высших отделов ЦНС на деятельность скелетных мышц. Продолговатый мозг и мост Непосредственным продолжением спинного мозга у всех позвоночных животных и человека является продолговатый мозг. Продолговатый мозг и варолиев мост (мост мозга) объединяют под общим названием заднего мозга. Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов. В продолговатом мозге по сравнению со спинным мозгом нет четкого сегментарного распределения серого и белого вещества. Скопление нервных клеток приводит к образованию ядер, являющихся центрами более или менее сложных рефлексов. Из 12 пар черепных нервов, связывающих головной мозг с периферией организма - его рецепторами и эффекторами, восемь пар (V -ХII) берут свое начало в заднем мозге. В продолговатом мозгу и мосте расположены ядра следующих черепных нервов: пара V черепных нервов – тройничный нерв, имеет двигательное и чувствительное ядра. Двигательное ядро расположено в мосту, иннервирует жевательные мышцы и вызывает движения нижней челюсти а также напрягает мягкое небо и барабанную перепонку. Чувствительные ядра (среднемозговое, мостовое, спинальное) получают от кожи, слизистых оболочек, органов лица и головы тактильную, температурную, висцеральную, проприоцептивную, болевую импульсацию, входят в проводниковый отдел соответствующих анализаторов и участвуют в различных рефлексах (например, жевательном, глотательном, чихательном). пара VI черепных нервов – отводящий нерв; я дро отводящего нерва расположено в мосту. Иннервирует наружную прямую мышцу глаза, вызывает поворот его кнаружи. пара VII черепных нервов – лицевой нерв; я дра лицевого нерва находятся в мосту. Двигательное ядро вызывает сокращения мимической и вспомогательной жевательной мускулатуры, регулирует передачу звуковых колебаний в среднем ухе в результате сокращения стременной мышцы. Чувствительное ядро одиночного пути, иннервируя вкусовые луковицы передних 2/3 языка, анализирует вкусовую чувствительность, участвует в моторных и секреторных пищеварительных рефлексах. Верхнее слюноотделительное (парасимпатическое) ядро стимулирует выделение секретов подъязычной, подчелюстной слюнных и слезной желез. пара VIII черепных нервов — преддверно-улитковый нерв; его чувствительные ядра расположены в продолговатом мозге. Вестибулярные ядра, иннервируя рецепторы вестибулярного аппарата, участвуют в регуляции позы и равновесия тела (статические и статокинетические рефлексы), в вестибулоглазных и вестибуловегетативных рефлексах, входят в проводниковый отдел вестибулярного анализатора. Улитковые ядра, иннервирующие слуховые рецепторы, участвуют в слуховом ориентировочном рефлексе, входят в проводниковый отдел слухового анализатора. пара IX — языкоглоточный нерв; ядра языкоглоточного нерва расположены в продолговатом мозге. Двойное (двигательное) ядро вызывает поднимание глотки и гортани, опускание мягкого неба и надгортанника при глотательном рефлексе. Чувствительное ядро одиночного пути получает вкусовую, тактильную, температурную, болевую и интероцептивную чувствительность от слизистой оболочки глотки, задней трети языка, барабанной полости и каротидного тельца, входит в состав соответствующих анализаторов, участвует в рефлексах жевания, глотания, в секреторных и моторных пищеварительных рефлексах, в сосудистых и сердечных рефлексах (из каротидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секрецию околоушной слюнной железы. пара X — блуждающий нерв; ядра блуждающего нерва расположены в продолговатом мозге. Двойное (двигательное) ядро, иннервируя мышцы неба, глотки, гортани, участвует в рефлекса глотания, рвоты, чиханья, кашля, в формировании голоса. Чувствительное ядро одиночного пути, иннервируя слизистую оболочку неба, корня языка, дыхательных путей, аортальное тельце, органы шеи, грудной, брюшной полостей, участвует в качестве афферентного звена в глотательном, жевательном дыхательных, висцеральных рефлексах. Оно входит в проводниковый отдел интероцептивного, вкусового, тактильного, температурного и болевого анализаторов. Заднее (парасимпатическое) ядро, иннервируя сердце, гладкие мышцы и железы органов шеи грудной и брюшной полостей, участвует сердечных, легочных, бронхиальных, пищи верительных рефлексах. пара XI — добавочный нерв; двигательное ядро добавочного нерв расположено в продолговатом и спинном мозге. Иннервируя грудино-ключично-сосцевидную и трапециевидную мышцы, оно вызывает наклон головы набок с поворотом лица в противоположную сторону, поднимание плечевого пояса вверх приведение лопаток к позвоночнику. пара XII — подъязычный нерв; двигательное ядро подъязычного нерва расположено в продолговатом мозге; иннервируя мышцы языка, вызывает его движение в рефлексах жевания, сосания, глотания, в осуществлении речи. Таким образом, с участием ядер черепных нервов реализуется сенсорная и рефлекторная (соматическая и вегетативная) функции ствола мозга. Сенсорные функции. Продолговатый мозг регулирует ряд сенсорных функций: · рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; · первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; · рецепцию слуховых раздражений — в ядре улиткового нерва; · рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолговатого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения. Проводниковые функции. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечно-сосудистой системы. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений. Рефлекторные функции. В продолговатом мозге находятся центры как относительно простых, так и более сложных рефлексов. За счет продолговатого мозга осуществляются: · защитные рефлексы (мигание, слезоотделение, чиханье, кашлевой и рефлекс акта рвоты); Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга. Отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания. · установочные рефлексы, обеспечивающие тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов; Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо. · лабиринтные рефлексы, способствующие правильному распределению мышечного тонуса между отдельными группами мышц при изменении позы тела; Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы соответствующей моменту прямолинейного или вращательного движения.. · рефлексы, связанные с функциями систем дыхания, кровообращения, пищеварения. Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов. Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов. Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени. В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез. Рефлекторные центры продолговатого мозга. В продолговатом мозге располагается ряд жизненно важных центров. В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей. Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатого мозга и разделен на две части, вдоха и выдоха. Сосудодвигательный центр (регуляции сосудистого тонуса) функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса. Средний мозг. К образованиям среднего мозга относят ножки мозга, ядра III (глазодвигательный) и IV (блоковый) пар черепных нервов, четверохолмие, красные ядра и черное вещество (черную субстанцию). В ножках мозга проходят восходящие и нисходящие нервные пути. В строении среднего мозга полностью утрачиваются сегментарные признаки. В среднем мозге клеточные элементы образуют сложные скопления в виде ядер. Ядерные образования относятся непосредственно к среднему мозгу, а также к входящей в его состав ретикулярной формации. Передние бугры четверохолмия получают импульсы от сетчатой оболочки глаз. В ответ на эти сигналы осуществляется регуляция просвета зрачка и аккомодация глаза. Задние бугры четверохолмия получают импульсы от ядер слуховых нервов, расположенных в продолговатом мозге. Благодаря этому происходит рефлекторная регуляция тонуса мышц среднего уха, а у животных - поворот ушной раковины к источнику звука. Таким образом, при участии передних и задних бугров четверохолмия осуществляются установочные, ориентировочные рефлекторные реакции на световые и звуковые раздражения (движения глаз, поворот головы и даже туловища в сторону светового или звукового раздражителя). При разрушении ядер четверохолмия зрение и слух сохраняются, но отсутствуют ориентировочные реакции на свет и звук. С деятельностью бугров четверохолмия тесно связана функция ядер III и IV пар черепных нервов, возбуждение которых определяет движение глаз вверх, вниз, в стороны, а также сведение (конвергенция) и разведение глазных осей при переносе взора с удаленных предметов на близкие и обратно. Красные ядра участвуют в регуляции мышечного тонуса и в проявлении установочных рефлексов, обеспечивающих сохранение правильного положения тела в пространстве. При отделении заднего мозга от среднего тонус мышц-разгибателей повышается, конечности животного напрягаются и вытягиваются, голова запрокидывается. Следовательно, у здорового животного и человека красные ядра несколько притормаживают тонус мышц-разгибателей. Черное вещество также регулирует мышечный тонус и поддержание позы, участвует в регуляции актов жевания, глотания, кровяного давления и дыхания, т. е. деятельность черного вещества, как и красных ядер, тесно связана с работой продолговатого мозга. Таким образом, средний мозг регулирует тонус мышц, соответствующим образом его распределяет, что является необходимым условием координированных движений; Средний мозг регулирует ряд вегетативных функций организма (жевание, глотание, кровяное давление, дыхание). За счет среднего мозга расширяется, становятся многообразнее рефлекторная деятельность организма (ориентировочные рефлексы на звуковые и зрительные раздражения).   ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
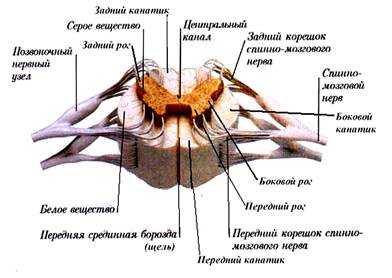 Внутри спинной мозг состоит из серого и белого вещества. Серое вещество заложено внутри и со всех сторон окружено белым. В каждой из половин спинного мозга оно образует два неправильной формы вертикальных тяжа с передними и задними выступами - столбами, соединенных перемычкой - центральным промежуточным веществом, в середине которого заложен центральный канал, проходящий вдоль спинного мозга и содержащий спинномозговую жидкость. В грудном и верхнем поясничном отделах имеются также боковые выступы серого вещества. Таким образом, в спинном мозге различают три парных столба серого вещества: передний, боковой и задний, которые на поперечном разрезе спинного мозга носят название переднего, бокового и заднего рогов.
Внутри спинной мозг состоит из серого и белого вещества. Серое вещество заложено внутри и со всех сторон окружено белым. В каждой из половин спинного мозга оно образует два неправильной формы вертикальных тяжа с передними и задними выступами - столбами, соединенных перемычкой - центральным промежуточным веществом, в середине которого заложен центральный канал, проходящий вдоль спинного мозга и содержащий спинномозговую жидкость. В грудном и верхнем поясничном отделах имеются также боковые выступы серого вещества. Таким образом, в спинном мозге различают три парных столба серого вещества: передний, боковой и задний, которые на поперечном разрезе спинного мозга носят название переднего, бокового и заднего рогов.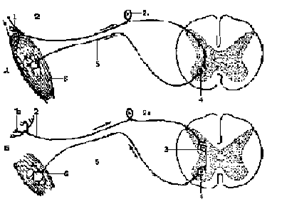 К числу рефлексов спинного мозга относятся защитные рефлексы, рефлексы на растяжение, мышц-антагонистов, висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, она указывает только на многообразие рефлекторных ответов. Даже у спинального животного трудно встретить рефлексы, которые относились бы только к одной из названных групп.
К числу рефлексов спинного мозга относятся защитные рефлексы, рефлексы на растяжение, мышц-антагонистов, висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, она указывает только на многообразие рефлекторных ответов. Даже у спинального животного трудно встретить рефлексы, которые относились бы только к одной из названных групп.