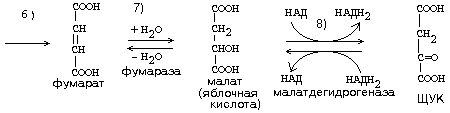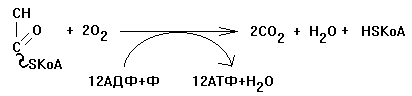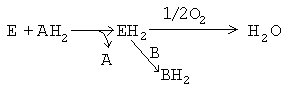|
|
ИСТОРИЯ РАЗВИТИЯ УЧЕНИЯ О БИООКИСЛЕНИИ.Стр 1 из 5Следующая ⇒ БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ. ПОНЯТИЕ О МЕТАБОЛИЗМЕ. Метаболизм – это совокупность химических реакций, протекающих в организме. При этом процессы, происходящие в просвете желудочно-кишечного тракта, не входят в понятие метаболизма, поскольку полость желудочно-кишечного тракта рассматривается как часть внешней среды. Метаболизм включает в себя более чем 100 000 разнообразных реакций, но существуют основные метаболические пути, построенные по единому плану. Такие пути могут быть линейными и разветвленными. Ферменты, катализирующие реакции, протекающие на этих путях, в организме объединены в мультиферментные системы. В мультиферментных системах продукт предыдущей реакции является субстратом для последующей. Метаболизм – это двуединый процесс, складывающийся из 2-х частей: катаболизма и анаболизма. В ходе катаболизма происходит разрушение, расщепление сложных веществ до более простых. В процессе анаболизма организм синтезирует собственные сложные органические вещества из простых. Оба процесса связаны между собой большим числом реакций, хотя в клетке часто бывают пространственно разделены. Однако, существуют химические реакции из числа обратимых, которые в равной степени можно отнести как к катаболизму, так и анаболизму. Принадлежность той или иной реакции к одному из этих процессов определяется тем, в какую сторону сдвинуто ее равновесие в данный момент времени. СХЕМА ЭТАПОВ КАТАБОЛИЗМА
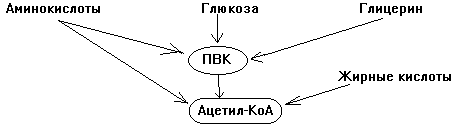
1-й этап. Образование мономеров из полимеров. Полимеры -------->Мономеры Белки ----------->Аминокислоты Крахмал --------->глюкоза Жиры ------------>глицерин + жирные кислоты
2-й этап. Превращение мономеров в ПВК и Ацетил-КоА.
3-й этап. Превращение Ацетил-КоА в конечные продукты катаболизма: СО2 и Н2О. Для всех классов веществ последний этап катаболизма одинаков: на 3-м этапе образуется большинство субстратов митохондриального окисления - 4 вещества из 9 основных и 5-й субстрат - ПВК. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ - это совокупность окислительных процессов в живом организме, протекающих с обязательным участием кислорода. Синоним - ТКАНЕВОЕ ДЫХАНИЕ. Окисление одного вещества невозможно без восстановления другого вещества. Окислительно-восстановительных процессов в живой природе очень много. Часть окислительно-восстановительных процессов, протекающих с участием кислорода, относится к биологическому окислению.
СОВРЕМЕННАЯ ТЕОРИЯ БИООКИСЛЕНИЯ Согласно СОВРЕМЕННОЙ ТЕОРИИ БИООКИСЛЕНИЯ в нашем организме окисление может происходить двумя способами: 1. Путем отнятия водорода от окисляемого субстрата: сюда относятся МИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ и ВНЕМИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ ОКСИДАЗНОГО ТИПА. 2. Путем присоединения кислорода к окисляемому субстрату - так происходит внемитохондриальное ОКИСЛЕНИЕ ОКСИГЕНАЗНОГО ТИПА (старое название - МИКРОСОМАЛЬНОЕ окисление).
ГЛАВНАЯ (ПОЛНАЯ) ЦЕПЬ 2) УКОРОЧЕННАЯ (СОКРАЩЕННАЯ) ЦЕПЬ 3) МАКСИМАЛЬНО УКОРОЧЕННАЯ (МАКСИМАЛЬНО СОКРАЩЕННАЯ) ЦЕПЬ.
Сначала разберем их строение на примере главной дыхательной цепи.
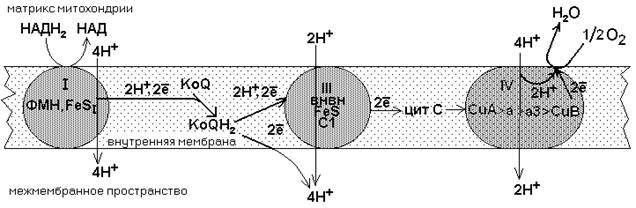
I. ГЛАВНАЯ ДЫХАТЕЛЬНАЯ ЦЕПЬ Главная дыхательная цепь - это три мультиферментных комплекса, встроенных во внутреннюю мембрану митохондрии. Обозначаются они латинскими цифрами – I, III и IV. СХЕМА ГЛАВНОЙ (ПОЛНОЙ) ДЫХАТЕЛЬНОЙ ЦЕПИ МИТОХОНДРИАЛЬНОГО ОКИСЛЕНИЯ
Комплекс I – НАДН-KoQ-редуктаза, комплекс III – KoQH2-редуктаза, комплекс IV – цитохромоксидаза. Есть еще комплекс II – сукцинат-KoQ-редуктаза, но он существует отдельно от остальных комплексов и не входит в состав главной цепи. Эти комплексы транспортируют водород от никотинамидных дегидрогеназ на кислород воздуха, в результате чего создается электрохимический градиент концентраций протонов - DmH+. Он возникает на внутренней мембране митохондрий между матриксом и межмембранным пространством. Его составляют два основных фактора: 1) Электрический мембранный потенциал Dy. 2) Градиент pH (осмотический или химический градиент). DmH+=Dy-k×DpH
DmH+ - положительная величина. Его можно выразить как в вольтах (V), так и в единицах энергии (кДж/моль). Изменение значения pH на одну единицу соответствует 0,06V или 5,7 кДж/моль. Энергия DmH+ используется для следующих процессов: Синтез АТФ. 2) Получение тепла (особенно важно для бурого жира и для мышечной ткани птиц). 3) Выполнение осмотической работы (транспорт фосфата в матрикс митохондрии). 4) Мышечная работа (в некоторых случаях). Для человека наиболее важен синтез АТФ. В полной цепи при окислении субстрата два атома водорода переносятся на НАД – кофермент никотинамидных дегидрогеназ.
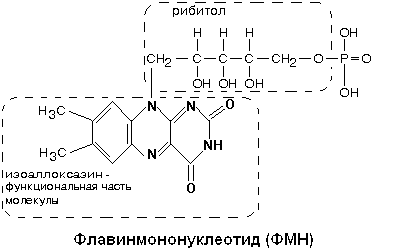
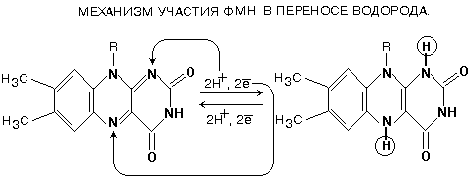
Как видно из приведенной схемы, в полной цепи при передаче двух атомов водорода на кислород воздуха, в межмембранном пространстве оказываются 10 протонов, перенесенных сюда из матрикса. Все переносчики встроены во внутреннюю мембрану митохондрий, кроме никотинамидных дегидрогенказ. Они составляют дыхательный ансамбль, тысячи таких ансамблей существуют в митохондрии и потребляют 90-95% кислорода, который используется клеткой. Два атома водорода отнимаются от субстрата и передаются на О2 с образованием Н2О. Разность потенциалов на двух концах полной цепи составляет 1.14V. КОМПЛЕКС I В составе комплекса находится 26 полипептидных цепей общей массой 800 кДа. Комплекс содержит следующие небелковые компоненты: Флавинмононуклеотид (ФМН), 5 центров FeS (железо-серные центры): FeS1a, FeS1b FeS2, FeS3, FeS4. В транспорте водорода по дыхательной цепи в этом комплексе принимает участие ФМН.
Одновременно с протонами транспортируются и электроны. Наибольшие перепады редокс-потенциала наблюдаются между железо-серными белками, расположенными в следующем порядке: ФМНÞFeS1aÞFeS1bÞFeS3ÞFeS4ÞFeS2 Комплекс I – интегральный белковый комплекс. Используя энергию, выделяющуюся при переносе электронов по дыхательной цепи, он транспортирует 4 протона из матрикса в межмембранное пространство – комплекс I работает как протонный генератор. Точный механизм этого транспорта до сих пор неизвестен. Далее комплекс I восстанавливает промежуточный переносчик KoQ (убихинон).
Это жирорастворимое низкомолекулярное вещество, содержащее длинную изопреновую цепь, не имеет белковой части. КоQ принимает водород от комплекса I. Образовавшийся КоQH2 отдает водород на комплекс III.
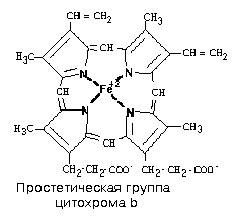
КОМПЛЕКС III. В своем составе содержит цитохромы – сложные белки, содержащие небелковый компонент - простетическую группу, сходню по строению с небелковой частью гемоглобина – гемом. 1) Цитохромы b, имеющие в своем составе два типа простетических групп тетрапиррольной структуры - «гем». Известно два гема цитохромов: be, обладающий низким окислительно-восстановительным потенциалом и bh с высоким окислительно-восстановительным потенциалом. Строение простетической группы цитохромов группы b, похожей на гем белка гемоглобина, представлено на рисунке. Его необходимо выучить.
3) Цитохром С1. Имеет в своем составе особый гем типа «с». Друг от друга цитохромы могут отличаться: 1) Строением белковой части; 2) Значением окислительно-восстановительного потенциала; 3) Строением радикалов, расположенных по периферии гема; 4) Присоединением гема к белковой части – в некоторых случаях гем присоединен к ней ковалентной связью за счет радикалов цистеина, что характерно для цитохромов c1 и c. От двух атомов водорода, которые переносятся на комплекс III от KoQ, дальше по цепи транспортируются только электроны, два протона (H+)комплекс III выбрасывает в межмембранное пространство вместе с еще одной парой протонов, которые подхватываются комплексом из матрикса. Таким образом, комплекс III в сумме выбрасывает в межмембранное пространство 4 протона. Поэтому комплекс III, как и комплекс I, является протонным генератором, и целью его работы также является создание DmH+.
КОМПЛЕКС IV. Комплекс IV называется цитохромоксидазой. Он способен захватывать из матрикса 4 протона. Два из них он отправляет в межмембранное пространство, а остальные передает на образование воды.
Благодаря многоступенчатой передаче энергия в дыхательной цепи выделяется не мгновенно, а постепенно (маленькими порциями) при каждой реакции переноса. Эти порции энергии не одинаковы по величине. Их величина определяется разницей между ОВП двух соседних переносчиков. Если эта разница небольшая, то энергии выделяется мало - она рассеивается в виде тепла. Но на нескольких стадиях ее достаточно, чтобы синтезировать макроэргические связи в молекуле АТФ. Такими стадиями являются: 1) НАД/ФАД - разность потенциалов 0.25V. 2) Цитохромы b/cc1 - 0.18V 3) aa3/O-2 - 0.53V. Значит, на каждую пару атомов водорода, отнятых от субстрата, возможен синтез 3-х молекул АТФ.
Макроэргическая связь - это такая ковалентная связь, при гидролизе которой выделяется не менее 30 кДж/моль энергии. Эта связь обозначается знаком ~. Синтез АТФ за счет энергии, которая выделяется в системе МтО, называется ОКИСЛИТЕЛЬНЫМ ФОСФОРИЛИРОВАНИЕМ. Основная роль АТФ - обеспечение энергией процесса синтеза АТФ. Для оценки эффективности работы системы МтО при окислении вычисляют КОЭФФИЦИЕНТ P/O. Он показывает, сколько молекул неорганического фосфата присоединилось к АДФ в расчете на один атом кислорода. Для главной (полная) цепи Р/О=3 (10H+/2H+ (затраты на освобождение АТФ из комплекса с ферментом) + 1H+ (затраты на транспорт фосфата) ) = 3,3 (округляют до 3-х)), коэффициент полезного действия системы - 65%, для укороченной P/O=2 (6H+/2H+ (затраты на освобождение АТФ из комплекса с ферментом) + 1H+ (затраты на транспорт фосфата) ) = 2, для максимально укороченной P/O=1 (4H+/2H+ (затраты на освобождение АТФ из комплекса с ферментом) + 1H+ (затраты на транспорт фосфата) ) = 1. Система МтО потребляет 90% кислорода, поступающего в клетку. При этом в сутки образуется 62 килограмма АТФ. Но в клетках организма содержится всего 20-30 граммов АТФ. Поэтому молекула АТФ в сутки гидролизуется и снова синтезируется в среднем 2500 раз (средняя продолжительность жизни молекулы АТФ - полминуты).
ОСНОВНЫЕ ПРОЦЕССЫ, ДЛЯ КОТОРЫХ ИСПОЛЬЗУЕТСЯ ЭНЕРГИЯ АТФ: 1. Синтез различных веществ. 2. Активный транспорт (транспорт веществ через мембрану против градиента их концентраций). 30% от общего количества расходуемого АТФ приходится на Na+,К+-АТФазу. 3. Механическое движение (мышечная работа).
СИНТЕЗ АТФ. Во внутренней мембране митохондрий расположен интегральный белковый комплекс – Н+-зависимая АТФ-синтаза seu Н+-зависимая АТФ-аза (два разных названия связаны с полной обратимостью катализируемой реакции), обладающий значительной молекулярной массой – более, чем 500кДа. Состоит из двух субъединиц: FO и F1. F1 представляет из себя грибовидный вырост на матриксной поверхности внутренней митохондриальной мембраны, FO же пронизывает эту мембрану насквозь. В толще FO расположен протонный канал, позволяющий протонам возвращаться обратно в матрикс по градиенту их концентраций. F1 способна связывать АДФ и фосфат на своей поверхности с образованием АТФ - без затраты энергии, но обязательно в комплексе с ферментом. Энергия необходима лишь для освобождения АТФ из этого комплекса. Эта энергия выделяется в результате тока протонов через протонный канал FO. В дыхательной цепи сопряжение абсолютно: ни одно вещество не может окисляться без восстановления другого вещества. Но при синтезе АТФ сопряжение одностороннее: окисление может идти без фосфорилирования, а фосфорилирование без окисления никогда не идёт. Это означает, что система МтО может работать без синтеза АТФ, но АТФ не может быть синтезирована, если не работает система МтО.
ВАРИАНТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ.
ПОЛНАЯ ДЫХАТЕЛЬНАЯ ЦЕПЬ В этой цепи окисляется небольшое количество субстратов, из которых главными являются четыре. Коэффициент Р/О=3.
Все ферменты полной цепи являются НАД-зависимыми дегидрогеназами.
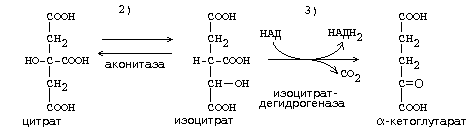
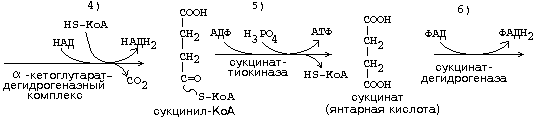
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦТК, лимоннокислый цикл, цикл Кребса) ЦТК, как и реакции митохондриального окисления, протекает в митохондриях. Представляет собой серию реакций, замкнутых в цикл.
Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь от образования цитрата до его превращения в ЩУК. В реакциях этого цикла участвуют четыре из девяти субстратов МтО. Происходит серия дегидрогеназных реакций. Из них 3-я, 4-я и 8-я происходят с участием НАД-зависимых дегидрогеназ, и каждая из этих реакций позволяет получить 3 молекулы АТФ. На 6-й стадии происходит ФАД-зависимая дегидрогеназная реакция, которая сопряжена с образованием 2-х молекул АТФ (Р/О = 2). На 5-й стадии 1 молекула АТФ образуется путем субстратного фосфорилирования. Итого за 1 оборот ЦТК образуется 12 молекул АТФ. Смысл ЦТК заключается в том, чтобы остатки уксусной кислоты расщепились с образованием большого количества АТФ. Кроме того, из ацетатных остатков образуется СО2 и Н2О, как конечные продукты обмена веществ. СО2 образуется в ходе ЦТК дважды: 1. на третьей стадии (окисление изоцитрата) 2. на четвертой стадии (окисление альфа-кетоглутарата). Если прибавить еще 1 молекулу СО2, которая образуется до начала ЦТК - при превращении ПВК в Ацетил-КоА, то можно говорить о трех молекулах СО2, образующихся при распаде ПВК. Суммарно эти молекулы, образующиеся при распаде ПВК, составляют до 90% углекислоты, которая выводится из организма. ИТОГОВОЕ УРАВНЕНИЕ ЦТК
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ЦТК
ГЛАВНАЯ РОЛЬ ЦТК - ОБРАЗОВАНИЕ БОЛЬШОГО КОЛИЧЕСТВА АТФ. 1. ЦТК - главный источник АТФ. Энергию для образования большого количества АТФ дает полный распад Ацетил-КоА до СО2 и Н2О. 2. ЦТК - это универсальный терминальный этап катаболизма веществ всех классов. 3. ЦТК играет важную роль в процессах анаболизма (промежуточные продукты ЦТК): - из цитрата -------> синтез жирных кислот - из aльфа-кетоглутарата и ЩУК ---------> синтез аминокислот - из ЩУК ----------> синтез углеводов - из сукцинил-КоА -----------> синтез гема гемоглобина
БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ. ПОНЯТИЕ О МЕТАБОЛИЗМЕ. Метаболизм – это совокупность химических реакций, протекающих в организме. При этом процессы, происходящие в просвете желудочно-кишечного тракта, не входят в понятие метаболизма, поскольку полость желудочно-кишечного тракта рассматривается как часть внешней среды. Метаболизм включает в себя более чем 100 000 разнообразных реакций, но существуют основные метаболические пути, построенные по единому плану. Такие пути могут быть линейными и разветвленными. Ферменты, катализирующие реакции, протекающие на этих путях, в организме объединены в мультиферментные системы. В мультиферментных системах продукт предыдущей реакции является субстратом для последующей. Метаболизм – это двуединый процесс, складывающийся из 2-х частей: катаболизма и анаболизма. В ходе катаболизма происходит разрушение, расщепление сложных веществ до более простых. В процессе анаболизма организм синтезирует собственные сложные органические вещества из простых. Оба процесса связаны между собой большим числом реакций, хотя в клетке часто бывают пространственно разделены. Однако, существуют химические реакции из числа обратимых, которые в равной степени можно отнести как к катаболизму, так и анаболизму. Принадлежность той или иной реакции к одному из этих процессов определяется тем, в какую сторону сдвинуто ее равновесие в данный момент времени. СХЕМА ЭТАПОВ КАТАБОЛИЗМА
1-й этап. Образование мономеров из полимеров. Полимеры -------->Мономеры Белки ----------->Аминокислоты Крахмал --------->глюкоза Жиры ------------>глицерин + жирные кислоты
2-й этап. Превращение мономеров в ПВК и Ацетил-КоА.
3-й этап. Превращение Ацетил-КоА в конечные продукты катаболизма: СО2 и Н2О. Для всех классов веществ последний этап катаболизма одинаков: на 3-м этапе образуется большинство субстратов митохондриального окисления - 4 вещества из 9 основных и 5-й субстрат - ПВК. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ - это совокупность окислительных процессов в живом организме, протекающих с обязательным участием кислорода. Синоним - ТКАНЕВОЕ ДЫХАНИЕ. Окисление одного вещества невозможно без восстановления другого вещества. Окислительно-восстановительных процессов в живой природе очень много. Часть окислительно-восстановительных процессов, протекающих с участием кислорода, относится к биологическому окислению.
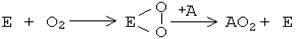
ИСТОРИЯ РАЗВИТИЯ УЧЕНИЯ О БИООКИСЛЕНИИ. А. Лавуазье в конце XVIII века показал, что животный организм потребляет из воздуха кислород и выделяет углекислый газ. Сделал вывод, что горение и окисление - это одно и то же, что биологическое окисление представляет собой "медленное горение", происходящее в присутствии воды и при низкой температуре. В конце XIX века русские исследователи А.Н. Бах и В.И.Палладин, работая независимо друг от друга, предложили 2 основные теории для объяснения процессов, протекающих в ходе биологического окисления. 1-я теория: А.Н.Бах (1857-1946) полагал, что в живых клетках существуют особые ферменты - "оксигеназы", которые взаимодействуют с кислородом, образуя перекиси. Сам кислород является не очень активным окислителем. Зато перекиси ("активный кислород") являются очень сильными окислителями и способны передавать кислород окисляемому веществу. Эта теория известна как "перекисная" или "теория активации кислорода".
2-я теория: В.И. Палладин (1859-1922) создал теорию "активации водорода". Считал, что универсальным путем окисления является отнятие от веществ (субстратов) водорода с участием специальных ферментов - хромогенов. После этого водород, по Палладину, может передаваться или на молекулу кислорода с образованием воды, или на другие молекулы, восстанавливая их.
Впоследствии теория В.И.Палладина блестяще подтвердилась для процессов митохондриального окисления, а ферменты, принимающие непосредственное участие в отнятии водорода от субстратов, в настоящее время называются дегидрогеназами.
  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
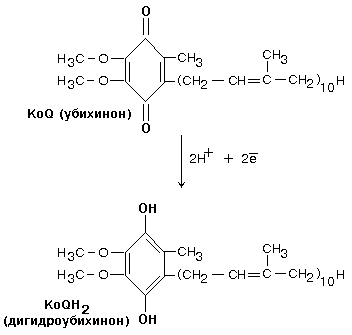
 2)FeSIII – железо-серный кластер.
2)FeSIII – железо-серный кластер.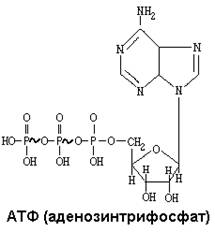 АДФ + Ф + ЭНЕРГИЯ -------> АТФ + Н2О
АДФ + Ф + ЭНЕРГИЯ -------> АТФ + Н2О