
|
|
Морфология и ультраструктура бактериальной клеткиОдин из основных признаков прокариотической клетки – отсутствие внутреннего разделения, обеспечиваемого элементарными мембранами. По сути, единственная мембранная система прокариот – ЦПМ, входящая в состав клеточной стенки, часто сложно устроенная и проникающая поперечными складками глубоко в цитоплазму.

Рис. 1. Строение прокариотической клетки Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с рибосомами и неоформленного ядра, называемого нуклеоидом. Могут быть дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили, включения. Некоторые бактерии способны образовывать споры для переживания неблагоприятных условий. Все структурные элементы бактериальной клетки условно можно разделить на обязательные (нуклеоид, ЦПМ, клеточная стенка, цитоплазма, 70S рибосомы) и необязательные, которые могут отсутствовать у некоторых видов (капсула, плазмиды, включения, жгутики, пили и др.) Капсула. Клеточную оболочку многих бактерий окружает слой аморфного, гидрофильного вещества. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов; например, капсула B.anthracis состоит из полимеров D-глутаминовой кислоты. Основную роль в организации капсул играет ЦПМ. Выделяют микрокапсулы (выявляемые только при электронной микроскопии в виде слоя мукополисахаридных фибрилл) и макрокапсулы (обнаруживают при световой микроскопии). У некоторых бактерий полимеры клеточной оболочки, выделяемые наружу, свободно располагаются вокруг нее, образуя слизистый слой. Слизь растворима в воде. Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Капсула и слизь предохраняют бактерии от повреждений, высыхания, фагоцитов, воздействия бактерицидных факторов, обеспечивают адгезию на различных субстратах, могут содержать запасы питательных веществ. Капсула антигенна (К-антиген). Для визуализации капсулы используют негативные методы окраски (по Бурри-Гинсу). Клеточная стенка. Клеточная стенка является уникальной, по своему химическому составу, структурой прокариотической клетки. Она выполняет ряд важных функций: защищает бактерии от внешних воздействий, придает им характерную форму, поддерживает постоянство внутренней среды, участвует в делении, на поверхности клеточной стенки находятся рецепторы для бактериофагов, бактериоцинов и различных химических веществ, через клеточную стенку осуществляется транспорт питательных веществ и выделение метаболитов. Это прочная, упругая структура, имеющая сложное строение. Опорным каркасом клеточной стенки бактерий является пептидогликан (муреин, мукопептид) – гетерополимер, состоящий из повторяющихся дисахаридных групп, соединенных поперечными и боковыми цепочками. Дисахарид, являющийся структурной единицей пептидогликана, состоит из N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных β-гликозидными связями. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки. Связывание фрагментов пептидогликана заключается в образовании пептидной связи между терминальным остатком аминокислотного мостика (D-аланином) с предпоследним остатком примыкающего аминокислотного мостика (L-лизином или диаминопимелиновой кислотой). Боковые мостики образуют четыре аминокислоты, поперечные (вертикально связывающие слои пептидогликана) – пять аминокислот. В состав боковых и поперечных мостиков входят уникальные аминокислоты, присутствующие только у прокариот, что делает эти связи мишенью для ряда антибактериальных препаратов.
    
   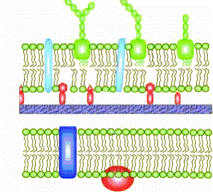
Рис. 2. Строение клеточной стенки бактерий В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Липополисахарид наружной мембраны состоит из 3 фрагментов: липида А — консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (от лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы. При нарушении синтеза клеточной стенки бактерий под влиянием различных факторов образуются клетки с измененной формой (чаще шаровидной): протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты — бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана и способные размножаться, называются L-формами. Существуют нестабильные L-формы, которые могут реверсировать, «возвращаясь» в исходную бактериальную клетку, и стабильные L-формы, не способные к реверсии. Цитоплазматическая мембрана. ЦПМ - это обязательная клеточная структура, являющаяся физическим, осмотическим, метаболическим барьером между внутренним содержимым бактериальной клетки и внешней средой. ЦПМ состоит из двух слоев фосфолипидов и встроенных в липидную мембрану белковых молекул (как большинство биологических мембран). Белки составляют 20-75%, липиды – 25-40% и в незначительных количествах в ЦПМ присутствуют углеводы и РНК, Белки ЦПМ подразделяют на структурные и функциональные. Первые образуют различные структуры ЦПМ, вторые представлены ферментами, участвующими в синтетических реакциях на поверхности мембраны и окислительно-восстановительных процессах, а также некоторыми специализированными энзимами (пермеазы участвуют в транспорте веществ). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Мезосомы образуют поперечные перегородки между делящимися клетками и являются местом прикрепления бактериальной хромосомы. У некоторых бактерий между ЦПМ и клеточной стенкой располагается периплазматическое пространство – полость шириной около 10 нм. Снаружи в это пространство открываются поры клеточной стенки, изнутри выходят некоторые клеточные ферменты (рибонуклеазы, фосфатазы, β-лактамазы). Цитоплазма. Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Ее основные составляющие – растворимые ферменты и РНК (мРНК и тРНК). Бактериальный геном. Эквивалентом ядра у бактерий является нуклеоид (генофор). Он расположен в центральной зоне бактерий в виде двойной суперспирализованной кольцевой молекулы ДНК. Она составляет 2-3% сухой массы клетки (более 10% по объему). Генофор не содержит гистонов. Объем генетической информации, кодируемой в бактериальной хромосоме зависит от вида бактерии. Кроме нуклеоида в бактериальной клетке имеются внехромосомные носители генетической информации – плазмиды, являющиеся ковалентно замкнутыми кольцами ДНК. Плазмиды несут ряд различных генов, кодирующих дополнительные (необязательные) признаки бактерий, например, гены антибиотикорезистентности (R-фактор). Бактерии могут обмениваться плазмидами в процессе конъюгации. Рибосомы. Бактериальные рибосомы – сложные глобулярные образования, состоящие из молекул РНК и связанных с ними белками. Рибосомы необходимы для синтеза полипептидов. Количество рибосом в различных бактериальных клетках колеблется от 5 до 50 тысяч. Диаметр рибосом составляет около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70S (единиц Сведберга), тогда как у эукариотических клеток 80S. Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50S и 30S (у эукариот 40S и 60S). Объединение субъединиц происходит перед началом трансляции. Рибосомальные РНК (рРНК) — консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субьединицы рибосом, а 23S рРНК — в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяющей оценить степень родства организмов. Различия в строении рибосом про- и эукариотических клеток делают рибосомы эукариот практически резистентными к действию антибиотиков, блокирующих синтез белка у бактерий. Запасные гранулы. В цитоплазме бактерий содержатся различные включения, которые содержат временный избыток метаболитов. В виде гранул могут запасаться полисахариды (крахмал, гликоген), жиры (триглицериды, запасаются у дрожжеподобных грибов рода Candida), полимеры β-оксимасляной кислоты, полифосфаты (волютин) у C.diphtheriae, сера, кристаллизованные белки и др. Жгутики. Жгутики бактерий являются органами движения (локомоции) бактерий. Расположение жгутиков – характерный признак, имеющий таксономическое значение. По количеству и расположению жгутиков различают монотрихи – один жгутик (V.cholerae), перитрихи (от греч. peri - вокруг и trichos - волос) – жгутики по всей поверхности бактериальной клетки (E.coli), лофотрихи (от греч. lophos - пучок и trichos - волос)– пучок жгутиков на одном конце клетки (Pseudomonas), амфитрихи (от греч. amphi - двойной, двусторонний и trichos - волос) –единичные жгутики или пучки жгутиков на разных полюсах клетки (Spirillum). Жгутик – спирально изогнутая полая нить, образованная субъединицами белка флагеллина. Толщина жгутиков 12-20 нм, длина 3-15 мкм (больше длины клетки). Жгутики состоят из 3 частей: спиралевидной нити, крюка и базального тельца. Базальное тельце содержит стержень со специальными дисками: одна пара дисков — у грамположительных и 2 пары дисков — у грамотрицательных бактерий. Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Жгутики являются H-антигенами, что используется в серологической идентификации. Подвижность бактерий определяют микроскопией препаратов «раздавленной» или «висячей» капли. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные – по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют ее). Микроворсинки. Помимо жгутиков, поверхность многих бактерий покрыта цитоплазматическими выростами – микроворсинками, встречающимися у подвижных и неподвижных видов. Пили (фимбрии, ворсинки) — нитевидные образования, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, прикрепление бактерий к поражаемой клетке; пили, ответственные за питание, водно-солевой обмен и половые (F-пили), или коньюгационные, пили. Пили многочисленны — несколько сотен на клетку. Однако половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col- плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях. Споры. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Способностью к спорообразованию обладают только бактерии родов Bacillus и Clostridium, включающие патогенные для человека виды, и многие сапрофиты. У бактерий рода Bacillus диаметр споры не превышает диаметр (поперечник) бактериальной клетки. У бактерий рода Clostridium (от лат. closter, веретено) диаметр споры превышает поперечник бактерии, что придает им форму веретена (C.perfringens), барабанной палочки (C.tetani) или теннисной ракетки (C.botulinum) при центральной, терминальной и субтерминальной локализации спор соответственно. Основное значение спор в выживаемости бактерий определяет их терморезистентность и резистентность к другим факторам (дезинфектанты и др.), оказывающим губительное воздействие на бактерии. Высокая устойчивость спор связана с низким содержанием свободной воды, высокой концентрацией кальция, вялостью метаболических процессов, наличием дипиколиновой кислоты и белка, богатого цистеином (что делает его похожим на кератин), а также наличие нескольких оболочек, которые являются дополнительной защитой от неблагоприятных внешних воздействий. При попадании спор в благоприятные условия они прорастают, проходя три последовательные стадии: активацию, инициацию, вырастание. Споры плохо окрашиваются и остаются бесцветными в окрашенных клетках при использовании обычных методов окраски (простые методы, метод Грама). Для окраски спор используют метод Ожешко (Ауэски). Основные формы бактерий Отдельным видам бактерий с достаточным постоянством присущи определенная форма, размер и расположение. Длина бактериальных клеток варьирует от 0.1-0,2 мкм (виды Mycoplasma) до 10-15 мкм (виды Clostridium), толщина – от 0,1 до 2,5 мкм. Средние размеры бактерий 2-3х0,3-0,8 мкм. Выделяют три основные формы бактерий – шаровидные (кокки), палочковидные (цилиндрические) и извитые (спиральные).
Рис. 3. Основные формы бактерий (микрофотографии) а) шаровидные (кокки); б) палочковидные; в) извитые Большинство кокков (от греч. kokkos - ягода, зерно) имеют шаровидную или овальную форму, клетки некоторых видов могут быть ланцетовидными (пневмококки) или бобовидными (нейссерии). По характеру расположения клеток в мазках выделяют диплококки (располагаются парами), стрептококки (располагаются цепочками), стафилококки (располагаются в виде гроздьев винограда), сарцины (располагаются пакетами по 8,16, 32 и более клеток) и др. Палочковидные бактерии различаются по форме концов клетки и взаимному расположению. Палочки могут быть правильной (кишечная палочка), неправильной (коринебактерии) формы, ветвящиеся (актиномицеты). Палочковидные бактерии могут располагаться в мазках одиночно и беспорядочно (монобактерии), попарно (диплобактерии) или цепочкой (стрептобактерии, стрептобациллы) (рис.4).
 
Рис. 4. Основные формы бактерий Извитые формы – представлены изогнутыми палочками. В зависимости от формы и количества завитков различают три типа клеток: вибрионы (от греч. vibrio - извиваюсь, изгибаюсь) имеют один завиток, не превышающий четверти оборота спирали (изогнутые клетки наподобие запятой); спириллы (от греч. speira – спираль) имеют 3–5 крупных завитков и спирохеты – большое количество мелких завитков (рис.5).

Рис. 5. Виды извитых клеток   Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом...  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|




 а)
а)  б)
б)  в)
в)