|
|
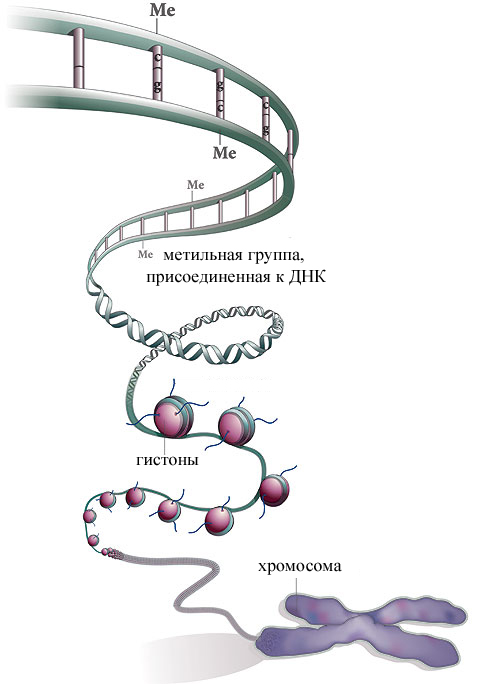
Какие механизмы и факторы приводят к изменению связи ДНК с гистонами.Гистоны Н2А, Н2В, Н3 и Н4 формируют октамерные структуры, вокруг которых закручивается нить ДНК, образуя таким образом нуклеосомы. Аминокислотые остатки гистонов могут подвергаться пост-трансляционным модификациям: Ацетилирование и деацетилирование гистонов -ацетилированию,- фосфорилированию,- метилированию. Ацетилирование связано с активацией транскрипции белки, осуществляющие ацетилирование - гистоновые ацетилтрансферазы (НАТ); донор ацетильной группы – ацетил коА; белки, осуществляющие деацетилирование – гистоновые деацетилазы (HDAC). Модель модификации гистонов: ДНК-связывающиеся активаторы привлекают НАТ для ацетилирования нуклеосомных гистонов, а репрессоры привлекают HDAC для деацетилирования гистонов. Эти события приводят к изменению структуры нуклеосом и активации или репрессии транскрипции соответственно. Эффект ацетилирования – ослабление связи между ДНК и гистонами из-за изменения заряда, в результате чего хроматин становится доступным для факторов транскрипции. Описать убиквитин зависимую протеосомную деградацию белков. Убиквитин-зависимая система протеолиза проводит поиск потенциальной мишени для протеолитической деградации среди внутриклеточных белков. Белки несут специфические сигналы деградации по аналогии с сигнальными последовательностями, которые направляют вновь синтезируемые белки к определенным микрокомпартментам клетки. Сигналы протеолитической деградации более сложные и разнообразные, так как с их помощью не только маркируются белки, удаляемые с помощью протеолиза, но и определяется время удаления и скорость их протеолитического расщепления. Для распознавания и декодирования таких сигналов в клетках эукариот имеется убиквитин-конъюгирующая система. Как в ядре, так и в цитоплазме эта система отделена пространственно и функционально от протеолитических ферментов, организованных в протеасомы. Распознанные данной системой белки-субстраты маркируются путем ковалентного присоединения к ним молекул стабильного 76- звенного белка - убиквитина. Убиквитин соединяется C-концом с боковыми остатками лизина в субстрате. Наличие такой метки в белке является первичным сигналом сортировки, направляющей образовавшиеся конъюгаты к протеасомам. В большинстве случаев к субстрату присоединяется несколько молекул убиквитина, которые организованы в виде бусинок на нитке. Молекулы белков, содержащие убиквитин, по-видимому, являются для протеасом предпочтительными субстратами. Конъюгацию убиквитина с субстратом можно представить следующим образом Убиквитин-активирующий фермент (E1) связывает убиквитин, гидролизует ATP и образует тиоэфирную связь между AMP и убиквитином с последующим переносом молекулы убиквитина на один из своих остатков Cys. Молекула активированного убиквитина далее соединяется с одним из ферментов семейства убиквитин-конъюгирующих ферментов (E2) и часто вслед за этим с убиквитин-лигазой (E3). Процесс конъюгации убиквитина с субстратом может катализироваться как самим E2, так и E2 совместно с E3. Белки E2 и E3 в клетках существуют в виде больших семейств, члены которых различаются по свойствам и внутриклеточной локализации. Мутации в генах семейства E2 у дрожжей показывают, что в ДНК-репарацию, прохождение клеточного цикла, биогенез пероксисом, в обеспечение устойчивости к тепловому шоку и ионам кадмия вовлечены разнообразные ферменты. Некоторые из ферментов E2 способны образовывать между собой гетеродимеры, которые в сочетании с белками E3 обеспечивают весь репертуар субстратных специфичностей убиквитин-конъюгирующих комплексов. Описать инициацию трансляции у прокариот: сигналы для инициации трансляции-последовательность Шайна-Дальгарно; трансляционные факторы, инициаторная т-РНК. (Я знаю можешь не читать. Читай следующий вопрос) Элонгация и терминация трансляции: функции факторов элонгации (EF-Tu, EF-Ts, EF-G), функции рибосомальных 3 центров связывания т-РНК, транслокация т-РНК, образование пептидной связи, два релизинговых фактора. Узнающие стоп кодоны. У эукариот имеется два фактора элонгации - eEF1 и eEF2. Не исключено, что в митохондриях и хлоропластах существуют независимые факторы элонгации, ответственные за связывание аминоацил-тРНК с рибосомой, но они еще не охарактеризованы (Льюин Б., 1987). Эукариотические клетки содержат в большом количестве фактор элонгации eEF1A, который является функциональным гомологом бактериального фактора EF-Tu. Так же как и у бактерий, этот фактор образует тройной комплекс с GTP и аминоацил-тРНК, обеспечивая вхождение последней в А-участокэлонгирующей рибосомы. Два других эукариотических фактора eEF1B и eEF2 резко отличаются от бактериальных функциональных аналогов EF1B(EF-Ts) и EF2(EF-G) по аминокислотным последовательностям. Гетеротримерный фактор eEF1B, как и его бактериальный аналог, катализирует обмен GDP на GTP в комплексе eEF1A-GDP. Фактор eEF2, по аналогии с бактериальными системами, обеспечивает транслокацию пептидил-тРНК в P-участок рибосом и перенос деацилированной тРНК в E-участок. У высших организмов этот фактор служит мишенью регуляторных воздействий через фосфорилирование. Замечательным свойством факторов eEF1A и eEF2 является способность связываться с компонентами цитоскелета эукариотических клеток. Полагают, что это их свойство может обеспечивать один из механизмов внутриклеточного транспорта мРНК, направляющих ее в полисомы.   Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|