|
|
Аллостерические ферменты; ковалентная модификация ферментов.Стр 1 из 7Следующая ⇒ Аллостерические ферменты; ковалентная модификация ферментов. Існують два принципових шляхи регуляції інтенсивності, або швидкості біохімічних ферментативних реакцій: А — через зміну каталітичної активності ферменту. Б — через зміну кількості ферменту (або ферментів), що визначають перебіг ферментативного процесу. А. Перший шлях регуляції передбачає наявність у ферментному пулі клітини спеціальних регуляторних ферментів, які містяться звичайно на головних, ключових ланках метаболізму. Цей шлях забезпечує термінову адаптацію ферментного апарату організму і реалізується протягом декількох секунд або хвилин —механізм “швидкого реагування”. Існують чотири основних механізми регуляції каталітичної активності ферментів 1. Алостерична регуляція активності ферментів. 2. Регуляція активності ферментів за рахунок їх ковалентної модифікації. 3. Активація ферментів шляхом обмеженого протеолізу. 4. Активація та гальмування активностей ферментів за допомогою особливих регуляторних білків. Алостеричні ферменти — це різновид регуляторних ферментів, що, крім активного центру, мають додатковий регуляторний (алостеричний) центр, з яким взаємодіють алостеричні регулятори (ефектори, модулятори). Алостеричні ефектори можуть бути як позитивними, тобто такими, що збільшують каталітичну активність ферменту (алостеричні активатори), так і негативними, тобто такими, що її гальмують (алостеричні інгібітори). За своєю молекулярною будовою алостеричні регуляторні ферменти складаються, як правило, з декількох поліпептидних ланцюгів, тобто мають четвертину структуру. Активний та регуляторний (алостеричний) центри локалізуються на різних білкових субодиницях — каталітичній та регуляторній, відповідно. Модифікація каталітичної активності такого ферменту здійснюється шляхом передачі на каталітичні субодиниці конформаційних змін із регуляторних субодиниць, які відбуваються в останніх після взаємодії з ліганда- ми — ефекторами. існують два фізичних стани алостеричного ферменту, що відрізняються своєю конформацією та каталітичною активністю: каталітичний (релаксований) стан (R-стан — relaxed, англ.) та інгібірований (напружений) стан (T-стан — tensed, англ.) Зворотний перехід між R- та T-станами залежить від взаємодії ферменту з алостеричними ефекторами (активатором або інгібітором, відповідно), які, взаємодіючи з місцями зв’язування на регуляторній субодиниці ферменту, стабілізують його молекулу в одному з конформаційних станів. Алостеричні ферменти каталізують біохімічні реакції, що знаходяться, як правило, на початку нерозгалужених або розгалужених метаболічних шляхів. При цьому модуляторами цих ферментів можуть бути як їх власні субстрати — гомотропні регуляторні ферменти, так і інші хімічні ефектори, зокрема кінцеві продукти багатоступеневого біохімічного процесу — гетеротропні регуляторні ферменти. прикладом ферменту з алостеричним механізмом регуляції є аспартаткарбамоїлтрансфераза (АКТ-аза) ЦТФ — кінцевий продукт біосинтезу — є алостеричним інгібітором АКТ-ази, що переводить фермент у неактивну Т-форму; АТФ — продукт біосинтезу пуринів та показник високого енергетичного стану клітини — є алостеричним активатором ферменту, що переводить його в активну R-форму. Ковалентна модифікація ферментів Постсинтетична ковалентна модифікація ферментних білків є одним із поширених механізмів контролю за перебігом метаболічних процесів. Шляхами такої модифікації є зворотне фосфорилювання-дефосфорилювання (найбільш поширений механізм регуляції), метилування, аденілування, АДФ-рибозилювання білків-ферментів. Фосфорилюють білки спеціальні ферменти протеїнкінази (протеїнфосфокінази), що за рахунок кінцевого (γ-) фосфату АТФ здійснюють фосфорилювання серинового чи треонінового (деякі протеїнкінази — тирозинового) радикалу відповідного білка: Зворотна реакція — дефосфорилювання білків — каталізується протеїнфос- фатазами: Субстратами протеїнкіназ є численні ферментні білки (глікогенфосфорилаза,кіназа фосфорилази b, глікогенсинтетаза, тригліцеридліпаза, піруватдегідрогеназа, ацетил-КоА-карбоксилаза тощо), деякі білки мембранних каналів, гістони хроматину тощоФосфорилювання багатьох білків-ферментів трансформує їх у каталітичноактивну форму (фосфорилювання глікогенфосфорилази, кінази фосфорилази b,тригліцеридліпази тощо); фосфорилювання інших ферментних білків (глікогенсинтетази, β-ГОМК-редуктази) є, навпаки, механізмом їх інактивації. Швидкість фосфорилювання-дефосфорилювання білків коливається, як правило, в межахвід декількох секунд до декількох хвилин, відповідно до необхідності включенняабо виключення певної фізіологічної функції клітини.У регуляції численних біохімічних функцій та фізіологічних процесів бере участьунікальна протеїнкіназа, що активується циклічним аденозинмонофосфатом(цАМФ) — вторинним посередником у дії на клітину багатьох фізіологічно активних сполук Циклические нуклеотиды (цАМФ, цГМФ) как регуляторы ферментативных Энзимопатии – врожденные (наследственные) дефекты метаболизма Углеводов, аминокислот, порфиринов, пуринов. Энзимотерапия – применение ферментов, их активаторов и ингибиторов в Медицине. Ензимотерапія, тобто використання ферментів і модуляторів (активаторів та інгібіторів) дії ферментів як лікарських засобів. До цих пір роботи в цьому напрямку майже не виходять за рамки експерименту. Виняток становлять деякі протеїнази: пепсин, трипсин, хімотрипсин і їх суміші (абомін, хімопсін), які застосовують для лікування ряду хвороб травного тракту. Крім протеїназ, ряд інших ферментів, зокрема РНКаза, ДНКаза, гіалуронідаза, колагенази, еластази, окремо або в суміші з протеиназа використовуються при опіках, для обробки ран, запальних вогнищ, усунення набряків, гематом, келоїдних рубців, кавернозних процесів при туберкульозі легень і ін Ферменти застосовуються також для лікування серцево-судинних захворювань, розчинення згустків крові. У нашій країні розроблено перший у світі препарат иммобилизованной стрептокінази, рекомендований для лікування інфаркту міокарда. Калікреїн-ферменти кінінової системи використовуються для зниження кров'яного тиску. Важливою і багатообіцяючою областю ензимотерапії є застосування інгібіторів ферментів. Так, природні інгібітори протеїназ (α 1-трипсин, α 2-хімотрипсин, α-макроглобулін) знайшли застосування в терапії гострих панкреатитів, артритів, алергійних захворювань, при яких відзначається активація протеолізу та фібринолізу, що супроводжується утворенням вазоактивних кінінів. Останнім часом набуло визнання застосування в онкологічній клініці ферментів бактеріальної природи в якості лікарських засобів. Принципы и методы выявления ферментов в биообъектах. Единицы измерения активности и количества ферментов. Обмен веществ (метаболизм) - общие закономерности протекания Энергетический баланс цикла трикарбоновых кислот. Физиологичное значение Реакций ЦТК. Біохімічний підсумок циклу трикарбонових кислот полягає в утворенні двох молекул СО2 (в ізоцитратдегідрогеназній та α-кетоглутаратдегідрогеназній реак-ціях) та чотирьох пар атомів водню, три з яких акцептуються НАД+ та одна— ФАД. Відновлені коферменти окислюються в дихальному ланцюзі мітохондрійутворюючи за рахунок окисного фосфорилювання по3 молекули АТФ на кожну молекулу НАДН і по2 молекули АТФ на кожну молекулу ФАДН2. Крім того, одна молекула АТФ утворюється в субстратному фосфорилюванні при перетворенні сукциніл-КоА в сукцинат. Таким чином, при повному окисленні однієї молекули ацетил-КоА до СО2 та Н2О в циклі трикарбонових кислот генерується12 молекул АТФ. 31. Реакции биологического окисления; типы реакций (дегидрогеназная, оксидазная, оксигеназная) и их биологическое значение. Тканевое дыхание. Цитохром Р-450 Ферментні системи, що каталізують реакції мікросомального окислення гідрофобних субстратів, є електронотранспортними ланцюгами, локалізованими в мембранах ендоплазматичного ретикулума гепатоцитів (та клітин деяких інших органів, що також беруть участь у реакціях детоксикації). Компонентами цих ферментних ланцюгів є ФАД-вмісний флавопротеїн, цитохром b5 та кінцева монооксигеназа — цитохром Р-450: Подібний цитохром Р-450-залежний електронотранспортний ланцюг каталізує реакції окислювального гідроксилювання стероїдів (синтезу та біотрансформації), що наявні в мітохондріях кори наднирникових та статевих залоз. Цитохром Р-450 — фермент, вперше відкритий у 1958 р. Американськими дослідниками Д.Гарфінкелем та М.Клінгенбергом (D.Garfinkel, M.Klingenberg). Це сімейство гемопротеїнів з молекулярною масою близько 50 кД; у різних біологічних об’єктах та тканинах виявлено більше 300 ізоформ цитохрому Р-450, що розрізняються за своєю субстратною специфічністю та особливостями первинної структури. Фізіологічне значення ізоформ цитохрому Р-450 полягає в захисті тваринного організму від численних низькомолекулярних ксенобіотиків, що надходять у внутрішнє середовище; ця система є додатковою до системи імунногозахисту (“друга імунна система”), яка протидіє надходженню в організм чужорідних високомолекулярних сполук біологічного походження. Біосинтез різних ізоформ цитохрому Р-450 кодується декількома сімействами генів, які у ссавців позначаються як CYP—гени (CYTOCHROMEP-450;англ.). У геномі людини за синтез цього гемопротеїну відповідають більше 100 генів, експресія яких призво- дить до продукції ізоформ із різною субстратною специфічністю (CYP1, CYP2, CYP3 тощо), що додатково поділяються на ензимні субсімейства.
38. Анаэробное окисление глюкозы, общая характеристика процесса. анаеробний гліколіз, що супроводжується утворенням з однієї молекули глюкози двох молекул молочної кислоти (лактату): 1. Активація молекули глюкози шляхом її фосфорилювання до фосфорного ефіру — глюкозо-6-фосфату. Ця реакція каталізується ферментом гексокіназою, що найбільш активна в м’язовійтканині
2. Перетворення (ізомеризація) глюкозо-6-фосфату у фруктозо-6-фосфат: Реакція каталізується ферментом фосфогексоізомеразою
3. Фосфорилювання фруктозо-6-фосфату з утворенням фруктозо-1,6-дифосфату. Джерелом фосфату, як і в 1-й реакції гліколізу, є молекула АTФ. Ферментом, що каталізує цю реакцію, є фосфофруктокіназа
4. Розщеплення фруктозо-1,6-дифосфату на дві молекули фосфотріоз шляхом розриву ковалентного –С–С– зв’язку між 3-м та 4-м атомами вуглецю в шестивуглецевому ланцюзі фруктозо-1,6-дифосфату. Реакція каталізується ферментом фруктозо-1,6-дифосфатальдолазою
5. Взаємоперетворення двох фосфотріоз (ДОАФ та Г-3-Ф), що каталізуєтьсяферментом тріозофосфатізомеразою:
6.1. Окислення гліцеральдегід-3-фосфату до 1,3-дифосфогліцерату (1,3-дифосфогліцеринової кислоти — 1,3-диФГК). Реакція каталізується ферментом гліцеральдегід-3-фосфатдегідрогеназою,
6.2. Перетворення 1,3-дифосфогліцерату на 3-фосфогліцерат (3-фосфоглі-церинову кислоту — 3-ФГК). Ця реакція супроводжується перенесенням макроергічної фосфатної групи від 1,3-диФГК на АДФ з утворенням молекули АТФ і каталізується ферментом фосфогліцераткіназою:
7. Перетворення 3-фосфогліцерату на 2-фосфогліцерат (2-фосфогліцериновукислоту — 2-ФГК). Реакція каталізується ферментом фосфогліцеромутазою:
8. Дегідратація 2-фосфогліцерату з утворенням фосфоенолпірувату (ФЕП). Реакція каталізується ферментом енолазою:
9. Утворення з фосфоенолпірувату пірувату. Реакція каталізується ферментом піруваткіназою
10відновлення пірувату до L-лактату:
39. Аэробное окисление глюкозы. Этапы превращения глюкозы до СО2 и Н2О. 1. Розщеплення глюкози до піровиноградної кислоти. Продуктом цього гліколітичного етапу розщеплення глюкози є піруват, а також двімолекули відновленого НАД+ і дві молекули АТФ:
2. Окислювальне декарбоксилювання піровиноградної кислоти. У результаті цього процесу утворюється ацетилкоензим А — основний субстратокислення в циклі трикарбонових кислот — та відновлена форма НАД+. Сумарне рівняння окислювального декарбоксилювання пірувату:
Окислювальне декарбоксилювання пірувату каталізується піруватдегідрогеназним комплексом — мультиферментною системою, яка в клітинах еукаріотів міститься вмембранах мітохондрій, а у прокаріотів — у цитоплазмі. До складу цього комплексувходять три ферменти, що каталізують три послідовні стадії перетворення піруватуна ацетил-КоА: піруватдегідрогеназа, дигідроліпоїлацетилтрансфераза, дигідроліпоїлдегідрогеназа та п’ять коферментів і простетичних груп: тіаміндифосфат(ТДФ), коензим А (КоА), ліпоєва кислота (ЛК), НАД+, ФАД. Б. У печінці. Глюкагон — стимулює глікогеноліз та гальмує глікогенез за механізмом,аналогічним дії адреналізу в клітинах м’язів. Інсулін — підвищує активність ферментативних реакцій синтезу глікогену зарахунок біохімічних механізмів, близьких до розглянутих вище. Таким чином, співвідношення глюкагон/інсулін є важливим фізіологічниммеханізмом, що контролює глікогенну функцію печінки та рівень глюкози в кровіпісля споживання їжі: – переважання інсуліну сприяє утворенню в організмі резервів вуглеводівуформі глікогену печінки; – переважання глюкагону сприяє мобілізації запасів глікогену печінки в умовахзниження рівня глюкоземії, яке спостерігається через кілька годин після споживання їжі.
46. Генетические нарушения метаболизма гликогена (гликогенозы, агликогенозы). Глікогенози Глікогенози — спадкові хвороби, молекулярною основою; уроджена недостатність синтезу певних ферментів глікогенолізу, пов’язана з дефектами в генетичній системі клітин. При глікогенозах у внутр органах татканинах (здебільшого в печінці, м’язах, клітинах крові) спостерігається накопичення аномально надмірної кількості глікогену, іноді зі зміненою молекулярноюструктурою, який не може використовуватися у метаболічних процесах —.Клінічно глікогенози проявляються важкою гіпоглюкоземією внаслідок нездатності глікогену печінки розщеплюватися з вивільненням молекул глюкози.Глікогенози, при яких ушкоджені ферментні системи мобілізації глікогену печінки, характеризуються збільшенням маси органа, жировою дистрофією гепатоцитівта явищами цирозу. Недостатність ферментних систем глікогенолізу в м’язах супроводжується судомами за умов фізичних. Аглікогенози — спадкові хвороби накопичення глікогену, порушення утворення ферменту глікогенсинтази - гепатоцити не здатні утворюватирезерви глікогену, концентрація якого всередині клітин значно зменшена. Внаслідок відсутності глікогенових резервів хворі при аглікогенозах, як і при глікогенозах, страждають від глибокої гіпоглюкоземії, особливо натщесерце, Гіпоглюкоземіям може супроводжуватися важкою комою енерг голодувголовного мозку. Такі хворі звичайно вмирають у ран дитвіці.
Ферменти глюконеогенезу піруваткарбоксилази:,карбоксибіотин,фосфоенолпіруваткарбоксикінази (ФЕП-кінази) Глюкозо-лактатний цикл (глюкозо-лактатний цикл, або цикл Корі), що пов’язує процеси утворення лактату в клітинах м’язової тканинив ході анаеробного гліколізу, його вихід у кров через плазматичні мембраним’язових клітин і використання лактату (після окислення в піруват) в гепатоцитахдля глюконеогенезу. Частина пірувату (не зазначена на схемі) окислюється в піруватдегідрогеназній реакції до ацетил-КоА.У скелетних м’язах ЛДГ-азна реакція перебігає переважно в бік утворення зпірувату лактату За рахунок відновлення пірувату в лактат таподальшого його використання для глюконеогенезу в печінці (цикл Корі), скелетнім’язи не тільки втрачають “зайву” молочну кислоту, що утворюється в особливозначних кількостях при інтенсивній фізичній праці, а й підтримують високеспіввідношення НАД+/НАДН, необхідне для активного функціонування гліколізу
Глюкозо-аланіновий цикл Важливим субстратом глюконеогенезу в печінці є аланін, який може утворюватися в скелетних м’язах у зворотній реакції трансамінування пірувату з глутаматом: піруват + глутамат-àL-аланін + a-кетоглутурат Вивільняючись із працюючих м’язів у кров, аланін поглинається гепатоцитами і (після перетворення в піруват)використовується в глюконеогенезі
49. Глюкоза крови (глюкоземия): нормогликемия, гипо- и гипергликемии,глюкозурия. Сахарный диабет – патология обмена глюкозы. 1. Зменшення рівня глюкоземії (гіпоглюкоземія, гіпоглікемія), що настає через декілька годин після останнього споживання їжі, супроводжується підвищенням рівня секреції a-клітинами острівкової частини підшлункової залозигормону глюкагону, який стимулює процеси глюконеогенезу за розглянутими вищемеханізмами. За рахунок активації каскадної аденілатциклазної системи в мембранах гепатоцитів глюкагон стимулює фосфороліз глікогену, що також робитьсвій внесок у збільшення рівня вільної глюкози. 2. Збільшення рівня глюкоземії (гіперглюкоземія, гіперглікемія) стимулюєсекрецію ß-клітинами підшлункової залози гормону інсуліну, який підвищує ступіньпроникності плазматичних мембран багатьох клітин для глюкози (окрім головногомозку), сприяючи їх внутрішньоклітинному метаболізму. Інсулін зменшує швидкість синтезу ферментів глюконеогенезу в печінці та, навпаки, стимулює синтезключових регуляторних ферментів гліколізу — гексокінази, фосфофруктокінази,піруваткінази, переводячи, таким чином, обмін глюкози з глюкогенного на гліколітичний шлях. Крім того, інсулін стимулює синтез у печінці та м’язах глікогену,що також є метаболічним процесом, спрямованим на зменшення концентраціївільної глюкози. Цукровий діабет розрізняють: – інсулінозалежний цукровий діабет (ІЗЦД), який розвивається внаслідокруйнування значної кількості (звичайно більше 90 %) секретуючих інсулін ß-клітин. Причиною деструкції ß-клітин є генетично зумовлений автоімунний процес.ІЗЦД складає 10-15 % всіх випадків цукрового діабету і проявляється гіперглікемією та схильністю до кетонемії та кетоацидозу. Ця форма ц дрозвивається звичайно в ранньому віці (до 30 років), найчастіше у дітей тапідлітків; – інсулінонезалежний цукровий діабет (ІНЗЦД) — форма цукрового діабету,за якого у більшості хворих зберігаються ß-клітини в інсулярній частиніпідшлункової залози, але порушені специфічні реакції клітин на дію інсуліну або регуляція його секреції під впливом збільшеної концентрації глюкози крові.ІНЗЦД розвивається звичайно у дорослих (старше 30 років) та осіб похилоговіку і проявляється гіперглікемією та ожирінням. Найбільш характерним клініко-біохімічним проявом цукрового діабету є збільшення (порівняно з нормою) рівня глюкози в крові в умовах натщесерце абопісля прийому їжі, який перевищує значення, характерні для фізіологічної, аліментарної гіперглюкоземії, досягаючи значень 500 мг % та більше. При легких формах захворювання гіперглюкоземія не спостерігається в постабсорбтивному станівиявляється лише заумов визначення толерантності організму до глюкози, якездійснюється шляхом “цукрового навантаження”. Энзимопатии их обмена. ПЕРЕТВОРЕННЯ фрукТОЗИ 1) Неспецеф гексокіназа – катал фосфорилювання різних гексоз 2) 2)специфічн фруктокіназа-катал фосфорилювання фруктози за 1-м вуглецев атамом 3) 3) Д-фруктозо-1-фосфат розщепл в печінці під дією альдолази на дві тріоди 4) Гліцеральдегід фосорилюється –утв. Гліцеральдегід-3-фосфат\\тріокіназа Спадкова непереносимість фруктози, ПРи генетично обумовленому дефекті фруктозо - 1 - фосфатальдолази. Симптоми виникають, коли в раціон додають фрукти, соки, сахарозу. Блювота, біль у животі, діарея, гіпоглікемія і навіть кома і судоми виникають через 30 хв після прийому їжі, що містить фруктозу. У маленьких дітей та підлітків, які продовжують приймати фруктозу, розвиваються хронічні порушення функцій печінки і нирок. ПЕРЕТВОРЕННЯ ГАЛАКТОЗИ 1) Фосфорилювання Д-галакт\\галактокіназа Д-галактоза + АТФ=Д-галакт-1-фосфат +АДФ 2)перетвор Д-галактоз-1-фосфат в епімерну молек Д-глюкоз-1 фосфат\\галактозо-1-фосфат-уридинтрансфераза Галактоземия спадковим дефектом будь-якого з трьох ферментів, що включають галактозу в метаболізм глюкози Галактоземия, викликана недостатністю галактозо - 1 - фосфатуріділтрансферази (ГАЛТ), Це захворювання проявляється дуже рано, і особливо небезпечно для дітей, так як основним джерелом вуглеводів для них служить материнське молоко, що містить лактозу. Ранні симптоми дефекту ГАЛТ: блювота, діарея, дегідратація, зменшення маси тіла, жовтяниця. У крові, сечі і тканинах підвищується концентрація галактози і галактозо - 1 - фосфату. У тканинах ока (в кришталику) галактоза відновлюється альдоредуктази з утворенням галактітола (дульціта). У цій реакції як донора водню іспользуетсяNADPH. до розвитку катаракти, яка спостерігається вже через кілька днів після народження. Виникають порушення функції печінки: гепатомегалія, жирова дистрофія. У нирках таких хворих також підвищена концентрація галактітола і галактозо - 1 - фосфату, що впливає на їх функції. Відзначають порушення в клітинах півкуль головного мозку і мозочка, у важких випадках - набряк мозку, затримку розумового розвитку, можливий летальний результат. Для галактоземії, викликаної дефектом галактокінази, теж характерна катаракта, але при цьому захворюванні, на відміну від дефекту ГАЛТ, не відзначають порушень функцій печінки, нирок, мозку. Найбільш важкі наслідки зниження активності ГАЛТ пов'язують з впливом галактозо - 1 - фосфату на активність інших ферментів, що беруть участь у вуглеводному Інсулін гальмує процес ліполізу та вивільнення жирних кислот. Інгібіруюча дія інсулінувідносно ліполізу в адипоцитах реалізується за рахунок двох біохімічних механізмів: а) зменшення концентрації цАМФ, що може бути пов’язаним з активацієюфосфодіестерази цАМФ; б) збільшення проникності мембран адипоцитів до глюкози, результатом чогоє активація в жировій тканині гліколізу і, відповідно, накопичення гліколітичнихметаболітів діоксіацетонфосфату та 3-фосфогліцеринальдегіду. Ці метаболіти, всвою чергу, є попередниками гліцерол-3-фосфату,
55. Реакции окисления жирных кислот (ß-окисление); роль карнитина втранспорте жирных кислот в митохондрии.
Окислення жирних кислот відбув в матриксі мітохондрій у результатіциклічного процесу, який включає в себе послідовне відщеплення від довголанцюгових молекул насичених жирних кислот, що складаються з парної кількостівуглецевих атомів (пальмітинової — С 16, стеаринової — С 18тощо), двовуглецевихфрагментів — циклу ß -окислення. Активація жирних кислот відбувається в цитоплазмі за участю специфічних фер- ментів ацил-КоА-синтетаз (тіокіназ), що утворюють КоА-похідні жирних кислот: R–COOH + KoA–SH + ATФ=R–CO–S–KoA + AMФ + ФФнЖирна Ацил-КоАкислота Ферментативні реакції ß - окислення жирних кислот 1. Дегідрування КоА-похідних жирних кислот за участю ФАД-залежного ферменту ацил-КоА-дегідрогенази. 2. Гідратація ненасиченого КоА-ацилу ферментом еноїл-КоА-гідратазою з утворенням спиртового похідного ацил-КоА — 3-оксіацилу-КоА (ß-гідроксіацилу-КоА) 3. Дегідрування оксипохідного ацил-КоА НАД-залежним ферментом 3-оксіацилКоА-дегідрогеназою. Продукт реакції — 3-кетоацил-КоА (ß-кетоацил-КоА): 4. Тіолітичне розщеплення 3-кетоацил-КоА за рахунок взаємодії з молекулоюКоА при участі ферменту ß- кетоацил-КоА-тіолази. В результаті реакції утворюється молекула КоА-похідного жирної кислоти, скороченого на два вуглецевіатоми, та ацетил-КоА: У результаті одного циклу ß-окислення змолекули жирної кислоти вивільняється однамолекула ацетил-КоА і, відповідно, вихіднамолекула ацил-КоА скорочується на два вуглецевих атоми. для повного розщеплення до ацетил-КоА будь-якоїмолекули жирної кислоти з парною кількістю вуглецевих атомів (n) потрібно (n/2 – 1)циклів ß-окислення.
Окислення гліцеролу Гліцерол, що утворюється при розщепленні триацилгліцеролів або гліцерофосфоліпідів, може вступати на шлях катаболізму (окислення) або знову використовуватися для біосинтезу різних класів гліцеридів 1. Включенню гліцеролудо метаболічних перетворень передує його активація,яка полягає в його трансформації за участю АТФ до гліцерол-3-фосфату (a- гліцерофосфату) при дії ферменту гліцеролфосфокінази. 2. a-Гліцерофосфат здатний до окислення мітохондріальним ферментомa гліцерофосфатдегідрогеназою з утворенням гліцеральдегід-3-фосфату (Г-3-Ф).Гліцеральдегід-3-фосфат є одним з центральних метаболітів гліколітичногоокислення глюкози. Подальше перетворення Г-3-Ф, утворенного при окисленнігліцеролу, співпадає з катаболізмом гліколітичного Г-3-Ф Г-3-Ф......1,3-диФГК......3-ФГК......2-ФГК......ФЕП......піруват Физиологическое значение
В нормі основ шляхомвикористання ацетил-КоА, (утв при ß-окисленні жирн кис), є цикл ТКРазом з тим, у печінці існує фізіол важл шлях утилізації ацетил-КоА,-- утвор молекул альтернативного метаболічного палива, яківикористовуються в інших тканинах —так званих кетонових (ацетонових) тілах. До кетонових тіл належать ацетоацетат, ß- гідроксибутират та ацетон.
2. Взаємодія ацетоацетил-КоА з молекулою ацетил-КоА з утворенням ß -гідрокси- ß метил-глутарил-КоА (ß-ГОМК).При утворенні кетоновихтіл реакція відбувається вмітохондріях і каталізуєтьсяферментом ß- ГОМК-синтетазою. 3. Розщеплення ß-гідрокси-ß-метилглутарилКоА мітохондріальнимферментом ß- ГОМК-ліазою з утворенням ацетоацетату та ацетил-КоА.ß -Гідроксибутират утворюється з ацетоацетату шляхом відновлення НАДзалежною ß -гідроксибутиратдегідрогеназою: Реакція перебігає в напрямку утворення ß-гідроксибутирату за умов високогоспіввідношення в гепатоцитах НАДН/НАД+, яке буває в умовах голодування. Ацетон утворюється в незначній кількості з ацетоацетату, що міститься в циркулюючій крові, за рахунок його неферментативного декарбоксилювання або діїферменту ацетоацетатдекарбоксилази. Ацетон видаляється з організму легенями; значне збільшення вмісту ацетону у видихуваному повітрі спостерігаєтьсяумовах декомпенсованого цукрового діабету. 65. .Із продуктами харчування до організму людини надходять ліпіди різних класів, а саме: триацилгліцероли (що складають основну масу харчових жирів); вільний холе- стерин та його ефіри з жирними кислотами (холестериди); складні ліпіди (переважно гліцерофосфоліпіди).Під дією панкреатичної ліпази та за участю жовчних кислот, які виробляються в печінці, триацилгліцероли продуктів харчування розщеплюються з утворенням 2-моноацилгліцеролів (моногліцеридів) та двох молекул вільних жирних кислот,що можна подати таким сумарним рівнянням:
Зазначені продукти гідролізу (вищі жирні кислоти, моногліцериди) абсорбу- ються клітинами слизової оболонки тонкої кишки (ентероцитами). Холестерин продуктів харчування всмоктується у вільному стану, холестериди — після відповідного гідролізу холестеролестеразою.Усередині ентероцитів продукти гідролізу триацилгліцеролів, що всмокталися, беруть участь у двох біохімічних процесах, які є передумовою подальшогонадходження нейтральних жирів у кров, біотранспорту та їх тканинного депонування, а саме: – реетерифікації вищих жирних кислот з утворенням нових молекул триацил- гліцеролів; – формування транспортних форм триацилгліцеролів — хіломікронів.Утворення хіломікронів. Триацилгліцероли ресинтезуються в ендоплазматичному ретикулумі мукозних клітин тонкої кишки. Вони утворюють ультрамікроскопічні краплинки, вкриті шаром поверхнево активних білків та фосфоліпідів. Ці структури отримали назву хіломікронів. До складу хіломікронів входять також вільний і етерифікований холестерин. Хіломікрони є основною молекулярною формою, у вигляді якої нейтральні жири (триацилгліцероли) проходять через латеральну мембрану ентероцитів і через систему лімфатичних судин (лактеалей) потрапляють у лімфатичний протік, а потім — у кров (через v.subclavia sin.). 66. Ліпопротеїни плазми крові Крім хіломікронів, кров людини містить декілька класів комплексів ліпідів із білками, що виконують функції міжорганного транспорту ліпідів — транспортніліпопротеїни плазми крові. Транспортні ліпопротеїни є фізико-хімічною формою, за допомогою якої гідрофобні молекули ліпідів утримуються в стабільному стані у гідрофільному (водно-сольовому) середовищі плазми крові. Фракціонування ліпопротеїнів крові людини здійснюється за допомогою методу ультрацентрифугування плазми в сольових розчинах, під час якого відбу- вається диференційоване спливання — флотація різних класів ліпопротеїнів, залежно від розмірів їх часточок та щільності. Ліпопротеїни можна також роз- ділити методом електрофорезу, при якому вони пересуваються разом із певними класами глобулінів (α-, β-ліпопротеїни тощо). За основу сучасної клініко-біохімічної класифікації ліпопротеїнів плазми крові людини взято їх розділення за умов ультрацентрифугування. Існує певна відпо- відність між класами ліпопротеїнів, що розділяються при застосуванні зазначених двох методів фракціонування. Основні класи ліпопротеїнів плазми крові: – хіломікрони (ХМ); – ліпопротеїни дуже низької щільності (ЛПДНЩ), або пре- β-ліпопротеїни; – ліпопротеїни проміжної щільності (ЛППЩ); – ліпопротеїни низької щільності (ЛПНЩ), або β-ліпопротеїни; – ліпопротеїни високої щільності (ЛПВЩ), або α-ліпопротеїни. Зазначені класи ліпопротеїнів розрізняються за своїми фізико-хімічними харак-теристиками, біохімічним (ліпідним, білковим) складом та фізіологічними функціями.
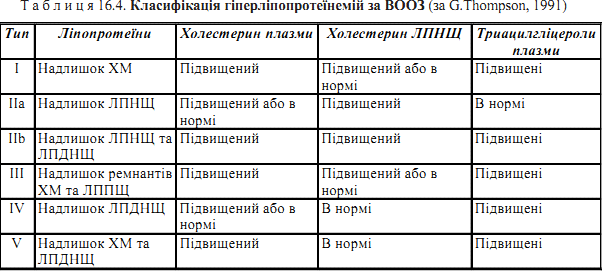
Хіломікрони — ліпопротеїни, що утворюються в слизовій оболонці тонкої киш- ки після внутрішньоклітинного ресинтезу триацилгліцеролів. Вони є молеку- лярною формою, в якій нейтральні жири та холестерин надходять із ентероцитів у кров через систему лімфатичних судин. ЛПДНЩ — ліпопротеїни, що також містять значну кількість нейтральних жирів. ЛПДНЩ синтезуються в гепатоцитах і є основною молекулярною формою, в якій триацилгліцероли виходять із печінки у кров та транспортуються в інші органи. ЛПНЩ — ліпопротеїни, що утворюються з ЛППЩ (ремнантів ЛПДНЩ).до складу ЛПНЩ входить найбільша кількість холестерину (здебільшого в етерифікованій формі), і вони є основним класом ліпопротеїнів плазми крові людини, що переносять холестерин ЛПВЩ — ліпопротеїни, що утворюються в печінці й, частково, у тонкій кишці Гіперліпопротеїнемія — клініко-біохімічний синдром, при якому в плазмі крові людини спостерігається підвищення (порівняно з нормою для певної попу- ляції) концентрації певних класів ліпопротеїнів, а також триацилгліцеролів та холестерину.
67. Атеросклероз — хвороба, головним проявом якої є відкладання в судинних стінках ліпідних утворень — “бляшок”, основними біохімічними компонентами яких є холестерин та його ефіри. Навкруги ліпідних бляшок в інтимі судин виникає клітинна реакція, що включає в себе утворення фіброзної тканини та проліфе- рацію гладенько-м’язових клітин. Атеросклеротичні бляшки спричиняють зву- ження кровоносних судин, посиленне згортання крові в ділянках їх локалізації та, як результат, порушення кровопостачання відповідних органів і тканин. Як наслідок атеросклерозу розвиваються ішемічна хвороба серця, інфаркт міокарда й порушення церебрального кровообігу, що стають важливою причиною смерті людей дорослого та похилого віку.Біохімічною основою розвитку атеросклерозу є підвищена концентрація в крові людини холестерину — гіперхолестеринемія, спричинена різними факторами — дієтарними, ендокринними, генетичними Ожиріння — стан, що характеризується надмірним накопиченням у жировій тканині триацилгліцеролів. При ожирінні збільшується кількість жирових клітин (адипоцитів) або їх розмір. Загальна маса нейтральних жирів в організмі людини за умов ожиріння може досягати значних кількостей. Порушення ліпідного обміну, що відбувається при ожирінні, часто поєднуються з наявністю у хворого атеросклерозу та/або цукрового діабету. Ожиріння розвивається внаслідок перевищення надходження та біосинтезу в тканинах нейтральних жирів (та інших біомолекул, які можуть перетворюватися в жири) над реальними енергетичними потребами організму в цих видах мета- болічного палива. Найбільш несприятливе значення для розвитку ожиріння має постійне надмірне надходження з продуктами харчування вуглеводів (особливо глюкози та фруктози) в кількостях, більших за ті, що безпосередньо окислюються в клітинах і можуть депонуватися у вигляді резервів глікогену. Цукровий діабет розглядається як патологія, що первинно пов’язана з порушеннями вуглеводного обміну. Дійсно, найбільш характернимбіохімічним проявом різних типів цукрового діабету в клініці є гіпергл   Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|