|
|
Шляхи біотрансформації холестерину. ⇐ ПредыдущаяСтр 7 из 7 Етерифікація холестерину Переважна частина холестерину тканин і близько 65 % холестерину плазми етерифіковано вищими жирними кислотами в положенні С-3. Синтез ефірів холестерину в плазмі крові та в клітинах відбувається за різними механізмами. 1. Зовнішньоклітинна етерифікація холестерину здійснюється ферментом лецитин (фосфатидилхолін)-холестерин-ацилтрансферазою (ЛХАТ) плазми. ЛХАТ каталізує реакцію перенесення ацильного залишку з 2-го (β-) положення фосфатидилхоліну на гідроксильну групу холестерину:
Біосинтез жовчних кислот. У гепатоцитах холестерин перетворюється на жовчні кислоти — важливі ком- поненти жовчі, що беруть участь у перетравленні харчових жирів у кишечнику людини і тварин.Жовчні кислоти є гідроксильоваими похідними холанової кислоти; до них належать такі сполуки: холева,дезоксихолева та літохолева кислоти. Першим етапом у біосинтезі жовчних кислот є 7 α-гідроксилювання холестерину, що каталізується ферментом мембран ендоплазматичного ретикулума 7 α-гідроксилазою, який єоднією з ізоформ цитохрому Р-450 та функціонує за участю НАДФН, кисню та вітаміну С (аскорбінової кислоти).7α-Гідроксилаза — регуляторний фермент, що є активним у фосфорильованій і малоактивним — в дефосфорильваній формі. Жовчні кислоти — кінцеві продукти цього метаболічногошляху — пригнічують активність ферменту за принципом негативного зворотного зв’язку. Після утворення 7 α-гідроксихолестеролу шлях біосинтезу жовчних кислот дихотомічно розгалужується: одна з гілок веде до утворення холевої кислоти, друга — хенодезоксихолевої. Ці сполуки надходять із гепатоцитів у жовчні капі- ляри і депонуюються в жовчному міхурі, надходячи з нього до порожнини дванад- цятипалої кишки. При дії ферментів мікроорганізмів, що містяться в кишечнику, утворюються вторинні жовчні кислоти — дезоксихолева та літохолева. Біосинтез стероїдних гормонів Стероїдні гормони містять у своєму складі 21 (кортикоїди, прогестерон) і менше (19 — андрогени, 18 — естрогени) атомів вуглецю, тому їх утворення з С27-стероїду холестерину включає, крім окисного гідроксилювання, і розщеплення вуглеводневого бічного ланцюга, реакції окислення, відновлення та ізомеризації.Першим етапом на шляху синтезу з холестерину стероїдних гормонів надниркових залоз (кортикостероїдів) є утворення С21 -стероїду прегненолону —безпосереднього попередника прогестагену прогестерону (С21), який у клітинахнадниркових залоз перетворюється на кортикостероїди (С21): глюкокортикоїд кортизол та мінералокортикоїд альдостерон.Гормони чоловічих та жіночих статевих залоз також утворюються з холестерину через стадію прегненолону та прогестерону, який у цих органах перетворюється в 17α-гідроксипрогестерон — попередник андрогену (С19) — тестостерону та естрогенів (С18) — естрону та естрадіолу. Біосинтез вітаміну D3 Перетворення холестерину у вітамін D3— холекальциферол — потребує розщеплення кільця циклопентанпергідрофенантрену з утворенням провітаміну D3, який підлягає реакціям окисного гідроксилювання з утворенням біологічно актив- ної форми вітаміну — 1,25-дигідроксихолекальциферолу (кальцитріолу) 65. .Із продуктами харчування до організму людини надходять ліпіди різних класів, а саме: триацилгліцероли (що складають основну масу харчових жирів); вільний холе- стерин та його ефіри з жирними кислотами (холестериди); складні ліпіди (переважно гліцерофосфоліпіди).Під дією панкреатичної ліпази та за участю жовчних кислот, які виробляються в печінці, триацилгліцероли продуктів харчування розщеплюються з утворенням 2-моноацилгліцеролів (моногліцеридів) та двох молекул вільних жирних кислот,що можна подати таким сумарним рівнянням:
Зазначені продукти гідролізу (вищі жирні кислоти, моногліцериди) абсорбу- ються клітинами слизової оболонки тонкої кишки (ентероцитами). Холестерин продуктів харчування всмоктується у вільному стану, холестериди — після відповідного гідролізу холестеролестеразою.Усередині ентероцитів продукти гідролізу триацилгліцеролів, що всмокталися, беруть участь у двох біохімічних процесах, які є передумовою подальшогонадходження нейтральних жирів у кров, біотранспорту та їх тканинного депонування, а саме: – реетерифікації вищих жирних кислот з утворенням нових молекул триацил- гліцеролів; – формування транспортних форм триацилгліцеролів — хіломікронів.Утворення хіломікронів. Триацилгліцероли ресинтезуються в ендоплазматичному ретикулумі мукозних клітин тонкої кишки. Вони утворюють ультрамікроскопічні краплинки, вкриті шаром поверхнево активних білків та фосфоліпідів. Ці структури отримали назву хіломікронів. До складу хіломікронів входять також вільний і етерифікований холестерин. Хіломікрони є основною молекулярною формою, у вигляді якої нейтральні жири (триацилгліцероли) проходять через латеральну мембрану ентероцитів і через систему лімфатичних судин (лактеалей) потрапляють у лімфатичний протік, а потім — у кров (через v.subclavia sin.). 66. Ліпопротеїни плазми крові Крім хіломікронів, кров людини містить декілька класів комплексів ліпідів із білками, що виконують функції міжорганного транспорту ліпідів — транспортніліпопротеїни плазми крові. Транспортні ліпопротеїни є фізико-хімічною формою, за допомогою якої гідрофобні молекули ліпідів утримуються в стабільному стані у гідрофільному (водно-сольовому) середовищі плазми крові. Фракціонування ліпопротеїнів крові людини здійснюється за допомогою методу ультрацентрифугування плазми в сольових розчинах, під час якого відбу- вається диференційоване спливання — флотація різних класів ліпопротеїнів, залежно від розмірів їх часточок та щільності. Ліпопротеїни можна також роз- ділити методом електрофорезу, при якому вони пересуваються разом із певними класами глобулінів (α-, β-ліпопротеїни тощо). За основу сучасної клініко-біохімічної класифікації ліпопротеїнів плазми крові людини взято їх розділення за умов ультрацентрифугування. Існує певна відпо- відність між класами ліпопротеїнів, що розділяються при застосуванні зазначених двох методів фракціонування. Основні класи ліпопротеїнів плазми крові: – хіломікрони (ХМ); – ліпопротеїни дуже низької щільності (ЛПДНЩ), або пре- β-ліпопротеїни; – ліпопротеїни проміжної щільності (ЛППЩ); – ліпопротеїни низької щільності (ЛПНЩ), або β-ліпопротеїни; – ліпопротеїни високої щільності (ЛПВЩ), або α-ліпопротеїни. Зазначені класи ліпопротеїнів розрізняються за своїми фізико-хімічними харак-теристиками, біохімічним (ліпідним, білковим) складом та фізіологічними функціями.
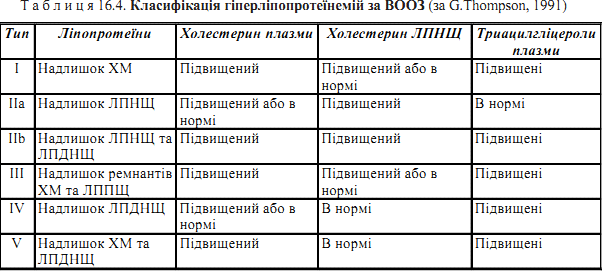
Хіломікрони — ліпопротеїни, що утворюються в слизовій оболонці тонкої киш- ки після внутрішньоклітинного ресинтезу триацилгліцеролів. Вони є молеку- лярною формою, в якій нейтральні жири та холестерин надходять із ентероцитів у кров через систему лімфатичних судин. ЛПДНЩ — ліпопротеїни, що також містять значну кількість нейтральних жирів. ЛПДНЩ синтезуються в гепатоцитах і є основною молекулярною формою, в якій триацилгліцероли виходять із печінки у кров та транспортуються в інші органи. ЛПНЩ — ліпопротеїни, що утворюються з ЛППЩ (ремнантів ЛПДНЩ).до складу ЛПНЩ входить найбільша кількість холестерину (здебільшого в етерифікованій формі), і вони є основним класом ліпопротеїнів плазми крові людини, що переносять холестерин ЛПВЩ — ліпопротеїни, що утворюються в печінці й, частково, у тонкій кишці Гіперліпопротеїнемія — клініко-біохімічний синдром, при якому в плазмі крові людини спостерігається підвищення (порівняно з нормою для певної попу- ляції) концентрації певних класів ліпопротеїнів, а також триацилгліцеролів та холестерину.
67. Атеросклероз — хвороба, головним проявом якої є відкладання в судинних стінках ліпідних утворень — “бляшок”, основними біохімічними компонентами яких є холестерин та його ефіри. Навкруги ліпідних бляшок в інтимі судин виникає клітинна реакція, що включає в себе утворення фіброзної тканини та проліфе- рацію гладенько-м’язових клітин. Атеросклеротичні бляшки спричиняють зву- ження кровоносних судин, посиленне згортання крові в ділянках їх локалізації та, як результат, порушення кровопостачання відповідних органів і тканин. Як наслідок атеросклерозу розвиваються ішемічна хвороба серця, інфаркт міокарда й порушення церебрального кровообігу, що стають важливою причиною смерті людей дорослого та похилого віку.Біохімічною основою розвитку атеросклерозу є підвищена концентрація в крові людини холестерину — гіперхолестеринемія, спричинена різними факторами — дієтарними, ендокринними, генетичними Ожиріння — стан, що характеризується надмірним накопиченням у жировій тканині триацилгліцеролів. При ожирінні збільшується кількість жирових клітин (адипоцитів) або їх розмір. Загальна маса нейтральних жирів в організмі людини за умов ожиріння може досягати значних кількостей. Порушення ліпідного обміну, що відбувається при ожирінні, часто поєднуються з наявністю у хворого атеросклерозу та/або цукрового діабету. Ожиріння розвивається внаслідок перевищення надходження та біосинтезу в тканинах нейтральних жирів (та інших біомолекул, які можуть перетворюватися в жири) над реальними енергетичними потребами організму в цих видах мета- болічного палива. Найбільш несприятливе значення для розвитку ожиріння має постійне надмірне надходження з продуктами харчування вуглеводів (особливо глюкози та фруктози) в кількостях, більших за ті, що безпосередньо окислюються в клітинах і можуть депонуватися у вигляді резервів глікогену. Цукровий діабет розглядається як патологія, що первинно пов’язана з порушеннями вуглеводного обміну. Дійсно, найбільш характернимбіохімічним проявом різних типів цукрового діабету в клініці є гіперглікемія (гіперглюкоземія), яка розвивається внаслідок втрати специфічного впливу інсуліну напроникність клітинних мембран для глюкози. Але метаболічні ефекти інсуліну розповсюджуються на багато аспектів обміну глюкози, ліпідів та амінокислот, у в’язку з чим цукровий діабет є хворобою, при якому відбуваються глибокі порушення не тільки вуглеводного, але й ліпідного та білкового обмінів. розрізняють інсулінозалежний цукровий діабет (діабет I типу, ювенільний діабет) та інсулінонезалежний цукровий діабет (діабетII типу, діабет похилого віку). Найбільш виражені порушення ліпідного обміну спостерігаються при діабеті II типу, який, як правило, поєднується з ожирінням. 68. Загальний пул амінокислот у тілі людини складається з потоків, які забезпе- чують надходження вільних амінокислот та їх використання в різноманітних анаболічних та катаболічних процесах.Потік амінокислот, що входить до амінокислотного пулу, складається з таких джерел: 1. Амінокислот, які всмоктуються ентероцитами кишечника внаслідок гід- ролізу харчових білків у травному каналі (шлунку, тонкому кишечнику); 60-100 г на добу. Додаткову компоненту в цей потік (від 35 до 200 г білка) вносить протеоліз ендогенних білків з епітелію ентероцитів, що злущується. 2. Амінокислот, які вивільняються в результаті розщеплення власних клітинних і позаклітинних білків. Середня тривалість напівжиття (Т1/2) білків становить від декількох годин для певних ферментів до декількох років для колагену. 3. Амінокислот, які синтезуються в організмі.До замінних амінокислот належать: аланін, аспарагінова кислота, аспарагін,глутамінова кислота, глутамін, пролін, гліцин, серин.
Потік амінокислот, що виходить з амінокислотного пулу, включає анабо- лічні і катаболічні шляхи перетворення вільних амінокислот і складається з таких компонентів: 1. Використання амінокислот длясинтезу білків організму. Для синтезу власних ферментних,структурних білків та фізіологічно активних сполук білкової і пептидної природи (на анаболічні потреби) використовується близько 75-80 % амінокислот, що ви- вільняються при розщепленні тканинних білків, та амінокислот, які надходять із кишечника. 2. Використання амінокислот, які не включені в анаболічні процеси, в катабо- лічних реакціях. При цьому молекули амінокислот розщеплюються з утворенням діоксиду вуглецю, води (через цикл лимонної кислоти) та кінцевих продуктів азотистого обміну (у людини — переважно сечовини). Певна частина безазотис- того вуглецевого скелета амінокислот використовується для утворення глюкози (глюконеогенезу) та кетонових тіл (кетогенезу). 69. ТРАНСАМІНУВАННЯ АМІНОКИСЛОТ
Реакції транс амінування полягають у переносі α-аміногрупи від амінокислоти на α-вуглецевий атом α-кетокислоти — акцептора аміногрупи (здебільшого — α-кетоглутарату); в результаті реакції утворюється α-кетоаналог вихідної амінокислоти та нова амінокислота (в разі використання як акцептора α-кетоглутарату — L-глутамат): Ферменти, що каналізують реакції трансамінування, — амінотрансферази (трансамінази).
Реакції трансамінування, що каталізуються амінотрансферазами, активно перебігають в печінці, скелетних м’язах, міокарді, головному мозку, нирках. Визначення активності аланінамінотрансферази (аланінової трансамінази — АлТ) та аспартатамінотрансферази (аспарагінової трансамінази — АсТ) широко застосовується з метою діагностики пошкоджень внутрішніх органів. Механізм дії амінотрансфераз Амінотрансферази є складними білками-ферментами, простетичною групою в яких є коферментні форми вітаміну В6 (піридоксину, піридоксолу) — піридоксальфосфат (ПАЛФ) та піридоксамінфосфат (ПАМФ), що утворюється з ПАЛФ у процесі переносу аміногрупи.
70.
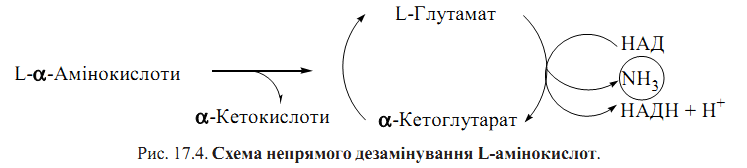
Пряме дезамінування Перший етап — утворення α-іміноглутарату — каталізується ферментом НАД- залежною глутаматдегідрогеназою, що локалізована в мітохондріях; другий етап — утворення α-кетоглутарату — є неферментативним. α-Кетоглутарат, що утворився, окислюється в циклі трикарбонових кислот, а аміак поглинається ферментативною системою синтезу сечовини. Зворотний процес — відновлювальне амінування α-кетоглутарату до L-глута- мату — може перебігати в цитозолі при участі цитозольної НАДФ-залежної глутаматдегідрогенази і бути допоміжним механізмом зв’язування аміаку. Непряме дезамінування - дезамінування вільних L-амінокислот за механізмом спряження реакцій трансамінування зα-кетоглутаратом і окислювального дезамінування L-глутамату
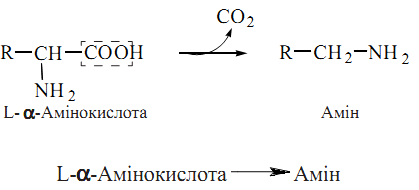
71. Реакція декарбоксилювання амінокислот полягає у відщепленні діоксину вуглецю від молекули амінокислоти з утворенням амінів (біогенних амінів), значна частина яких має високу фізіологічну активність як гормони, нейромедіатори, є їх попередниками або метаболітами:
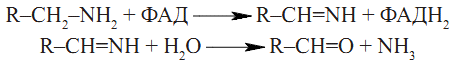
Фізіологічне значення декарбоксилювання амінокислот • Утворення фізіологічно активних сполук — гормонів, медіаторів. • Катаболізм амінокислот у процесі гниття білків у кишечнику. Окислення біогенних амінів Накопичення біогенних амінів в організмі спричиняє несприятливі патофізіологічні зміни з боку серцево-судинної системи, кишечника, інших гладеньком’язових органів. Знешкодження (детоксикація) фізіологічно активних амінів відбувається в клітинах печінки при участі моноамінооксидази мітохондрій — ФАД-залежного ферменту, що спричиняє окислювальне дезамінування амінів до альдегідів:
Альдегіди — продукти дезамінування біогенних амінів окислюються до відповідних кислот і підлягають подальшій окислювальній деградації або екскретуються з організму із сечею. Аміак надходить у систему синтезу сечовини.
72. Шляхи утворення аміаку в організмі людини 1. Головним у кількісному відношенні джерелом накопичення аміаку в організмі людини є окислювальне дезамінування амінокислот, тобто білковий катаболізм: азот сечовини — кінцевого азотовмісного продукту деградацію білків — складає близько 90 % всього азоту, що екскретується. Додатковими джерелами ендогенного аміаку є реакції дезамінування біогенних амінів, азотистих основ, які утворюються при катаболізмі нуклеотидів. 2. Утворення аміаку в головному мозку Основним джерелом утворення аміаку втканині головного мозку є реакція гідролітичного дезамінування АМФ до інозинмонофосфату (ІМФ), що каталізується аденозиндезаміназою:
Механізми знешкодження аміаку Залежно від молекулярної форми, у вигляді якої екскретуються кінцеві продукти азотистого(амінного) катаболізму, існує три типи тваринних організмів: 1) амоніотелічні організми — такі, що виводять амінний азот у вигляді розчин- ного іону амонію (до них належить більшість хребетних, що мешкають у воді); 2) урикотелічні організми — такі, що виводять амінний азот у вигляді сечової кислоти (птахи, наземні рептилії); 3) уреотелічні організми — основним продуктом знешкодження та екскрету- вання аміаку у яких є сечовина (більшість наземних хребетних, включаючи ссавців, зокрема організм людини). 73. Біосинтез сечовини синтез сечовини відбувається з аміаку та вугільної кислоти в результаті циклічного процесу, в якому каталітичну роль відіграють амінокислоти аргінін,орнітин та цитрулін (орнітиновий цикл Кребса-Хензелайта):
джерелами двох аміногруп, що використовуються для утворення молекули сечовини, є аміак, який вивільняється при окислювальному дезамінуванні L-глутамату, та аміногрупа амінокислоти L-аспартату. Генетичні дефекти ферментів синтезу сечовини Існують спадкові ензимопатії, спричинені повним або частковим дефектом утворення в печінці окремих ферментів циклу сечовиноутворення. Найбільш важкими клінічними проявами характеризуються порушення синтезу карбамоїлфосфатсинтетази та орнітинкарбамоїлтрансферази. Діти з такими генетичними дефектами страждають вираженою енцефалопатією, прояви якої дещо послаблюються в умовах повного виключення споживання харчових білків.
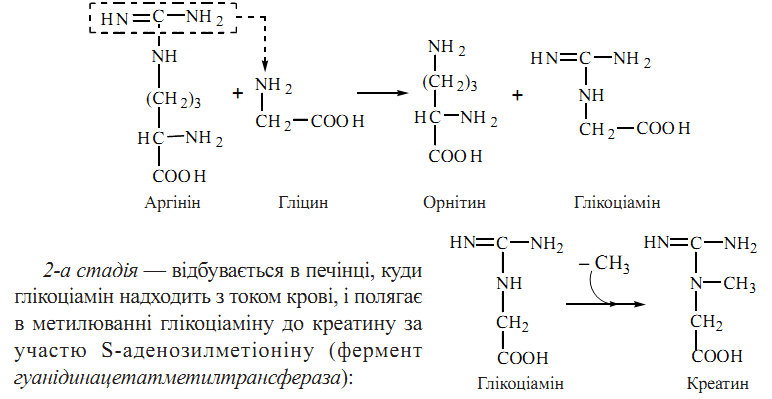
74. Глюкогенні амінокислоти L-Амінокислоти, що метаболізуються в циклі трикарбонових кислот можуть включати свої вуглецеві скелети в молекули глюкози. Ці амінокислоти,використання яких у синтезі глюкози реалізується після їх входження в ЦТК через ацетил-КоА, α-кетоглутарат, сукциніл-КоА та фумарат, отримали назву глюкогенних амінокислот. Кетогенні амінокислоти Дві L-амінокислоти включаються в катаболізм тільки через ацетоацетил-КоА, який у клітинах печінки може перетворюватися на кетонові тіла ацетоацетат та β-гідроксибутират. Це — кетогенні амінокислоти. Деякі амінокислоти віддають свої вуглецеві фрагменти на утворення як глюкози, так і кетонових тіл. Кетогенез із амінокислот має особливе негативне значення при деяких порушеннях ферментних процесів, зокрема при некомпенсованому цукровому діабеті,у зв’язку з чим таким хворим рекомендується обмежувати надходження кетогенних амінокислот у складі продуктів харчування. 75. Креатин — азотиста сполука, яка у вигляді креатинфосфату має важливе значення в енергозабезпеченні функції м’язів. Біосинтез креатину відбувається за участю амінокислот гліцину, аргініну та метіоніну. Процес синтезу складається з двох стадій: 1-ша стадія — відбувається в нирках і полягає в утворенні глікоціаміну (гуанідинацетату) із аргініну та гліцину (фермент гліцинамідинотрансфераза):
Фосфорилювання креатину при дії креатинфосфокінази генерує креатинфосфат — джерело термінової регенерації АТФ при м’язовому скороченні. Незворотна неферментативна дегідратація і дефосфорилювання креатин фосфату призводить до утворення ангідриду креатину — креатиніну.У формі креатиніну з організму людини виділяється із сечею значна частина азоту амінокислот; у здорової людини виділення креатиніну пропорційне масі м’язової тканин і значно збільшується за умов травматичних пошкоджень м’язів. 76. Глутатіон — трипептид γ-глутамініл-цистеїніл-гліцин, що має в своєму складі вільну сульфгідрильну групу:
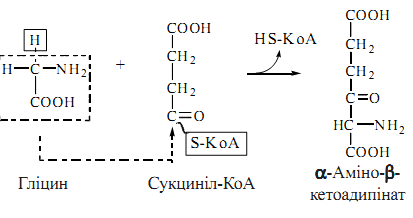
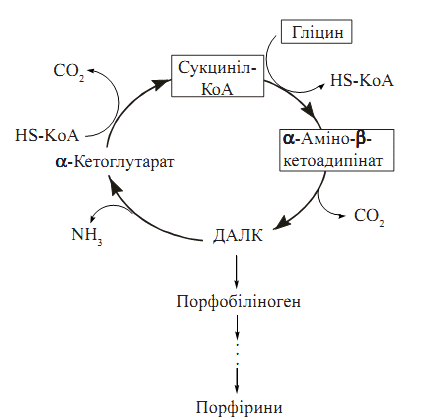
Глутатіон міститься в клітинах тваринного організму у високій концентрації. Глутатіон зворотно перетворюється з відновленої (Г–SH) до окисленої (Г–S–S–Г) форми, відіграючи роль буфера SH-груп.Біохімічна функція глутатіону в організмі пов’язана з відновленням і детоксикацією органічних пероксидів — похідних пероксиду водню НО–ОН, у молекулі якого один (гідропероксиди) або обидва атоми водню (алкілпероксиди) заміщені на алкільні радикали. 77. Шляхи метаболізму фенілаланіну 1.Катаболічний шлях обміну полягає у втраті фенілаланіном аміногрупи (в реакції трансамінування) з утворенням фенілпірувату та кінцевого метаболіту фенілацетату, що екскретується з організму. 2. Шлях синтезу фізіологічно активних сполук починається з перетворення фенілаланіну на тирозин при дії ферменту фенілаланінгідроксилази з подальшим перетворенням тирозину. Шляхи метаболізму тирозину 1. Катаболічний шлях обміну полягає у трансамінуванні тирозину і перетворенні на p-оксифенілпіруват, який окислюється до гомогентизинової кислоти у складній реакції, коферментну роль у якій виконує аскорбінова кислота(вітамін С); подальші перетворення полягають в окисленні гомогентизату до фумарилацетоацетату (фермент оксидаза гомогентизинової кислоти) та розщепленні фумарилацетоацетату до фумарату та ацетоацетату 2. Шлях синтезу катехоламінів та меланінів (пігментів шкіри). Шлях починається з окислення тирозину за участю специфічної гідроксилази до 3,4-діоксифенілаланіну (ДОФА), на рівні якого відбувається дивергенція двох обмінних шляхів: утворення катехоламінів (через декарбоксилювання до дофаміну) та меланінів (через окислення тирозиназою до дофахінону). 3. Шлях синтезу тиреоїдних гормонів — реалізується в клітинах щитовидної залози і полягає в утворенні йодованих тиронінів. 78. Фенілкетонурія — ензимопатія, спричинена генетичним дефектом синтезу фенілаланінгідроксилази. Внаслідок блокування утворення тирозину з фенілаланіну останній в збільшеній кількості надходить на шлях утворення фенілпірувату та фенілацетату, які в надмірних концентраціях накопичуються в організмі хворих. Патологія проявляє себе ранніми порушеннями психічного розвитку дитини —фенілпіровиноградна олігофренія. Алкаптонурія — ензимопатія, що викликана генетично детермінованою недостатністю ферменту оксидази гомогентизинової кислоти. Характерним проявом захворювання є надмірне виділення гомогентизинової кислоти із сечею, яка при додаванні лугів набуває темного забарвлення; акумуляція гомогентизату в тканинах суглобів призводить до розвитку артритів. Альбінізм — ензимопатія, біохімічною основою якої є спадкова недостатність ферменту тирозинази, що каталізує реакції, необхідні для утворення чорних пігментів меланінів. Відсутність меланінів у меланоцитах шкіри проявляється недостатньою (або відсутньою) пігментацією шкіри та волосся, підвищеною чутливістю шкіри до сонячного світла, порушенням зору. 79. Порфірини та їх комплекси з металами — металопорфірини — є простетичними групами багатьох гемопротеїнів — білків, які беруть участь в окислювально-відновлювальних реакціях у тваринних та рослинних клітинах. Представниками гемопротеїнів, що містять металопорфіринові групи, є Fe2+-вмісні гемоглобін і міоглобін (О2— запасаючий білок м’язів). Порфірини — сполуки циклічної будови, основою структури яких є ароматична гетероциклічна система — порфін. Порфін, в свою чергу, є тетрапіролом, який утворюється при сполученні між собою метенільними (–СН=) містками чотирьох кілець азотистого гетероциклу піролу. Синтез порфіринів
  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|