
|
|
Виды сенсорных модальностей, связанных с движением, и соответствующие им рецепторы(по Шеперду) точными движениями. Эти рецепторы представляют собой образования веретеновидной формы, заключенные в тонкую растяжимую соединительнотканную капсулу. Веретена располагаются в мышцах продольно и растягиваются при растяжении мышцы. Каждое веретено образовано несколькими волокнами (от 2 до 12), названными ин-трафузальными (от лат. fusus — веретено) (рис. 3.78). Эти волокна омывает тканевая жидкость. Интрафузальные волокна бывают двух типов. В центральной части большинства волокон располагается цепочка из одного ряда клеточных ядер. Второй тип волокон в центре несет ядерное скопление (волокна с ядерной сумкой); эти волокна длиннее и толще, чем первые. Периферические концы волокон обоих типов способны растягиваться. Интрафузальные волокна иннерви-руются афферентными миелинизированными нервными волокнами. При Рис. 3.78. Мышечные веретена: А — мышечные веретена в глазных мышцах: / — нервное волокно; 2 — мышечное волокно; 3 сухожилие; Б и Г — волокна с ядерной сумкой; йиД- волокна с цепочкой ядер в центре; [А, Б, В — гистологические препараты; Г, Д — схемы]; / — капсула; 2 — интрафузальные волокна; 3 — экстрафузальные волокна; 4 — первичные окончания; 5 — вторичные окончания; 6 — кустовидные эфферентные окончания; 7 — первичный афферент; 8 — вторичный афферент; 9 — эфферентные нервные волокна
этом толстое нервное волокно, имеющее большую скорость проведения импульсов, подходит к центральной части интрафузального волокна и по спирата обвивает ядерную сумку или область, содержащую цепочку ядер. Такое окончание называют первичным. По сторонам от первичных окончаний более тонкие афферентные волокна образуют вторичные окончания, форма которых может быть похожа на гроздь. Первичное окончание реагирует на степень и скорость растяжения мышцы, а вторичное — только на степень растяжения и изменение положения мышцы. При растяжении мышцы информация от нервных окончаний поступает в спинной мозг, где часть ее переключается на мотонейроны передних рогов. Их ответная рефлекторная импульсация приводит к сокращению мышцы. Другая часть импульсов переключается на вставочные нейроны и поступает в другие отделы нервной системы (см. ниже). Мышечные веретена имеют также и эфферентную иннервацию, которая контролирует степень их растяжения. Эфферентные волокна подходят к мышечным веретенам от мотонейронов спинного мозга, но не от тех, что иннервируют саму мышцу, волокна которой называют экстрафузалъными. Однако в некоторых случаях мышечные веретена получают моторную иннервацию по коллатералям от аксонов, идущим к мышцам. Это наблюдается, например, в мышцах глазного яблока. Кроме рецепторных окончаний, лежащих в самих мышцах и реагирующих на степень их растяжения, существуют рецепторы в местах соединения мышц с сухожилиями. Они носят название сухожильных органов (рецепторов) Голъджи (рис. 3.79). Они покрыты капсулой и иннервируются толстыми миелиновыми волокнами. Оболочка волокон теряется в месте прохождения через капсулу, и волокно образует терминальные ветвления между пучками коллагеновых волокон сухожилия. Эти окончания возбуждаются при сдавливании их волокнами сухожилия при сокращении мышцы, тогда как мышечные веретена неактивны, и наоборот, при растяжении мышцы активность веретен возрастает, а сухожильных рецепторов снижается. Большое количество рецепторных окончаний расположено в суставах (рис. 3.79). В суставных связках лежат рецепторы, схожие с сухожильными,
В Рис. 3.79. Рецепторные окончания в сухожилиях (по Догелю): / — миелиновое нервное волокно; 2 — мышечные волокна; 3 — концевое разветвление; 4 — пучок коллагеновых волокон, 5 — осевой цилиндр, 6 — ядро сухожильной клетки фиброцита; / — тельца Руффини; 8 — тельца Пачини; 9 — рецепторы Гольджи; 10 — свободные нервные окончания в соединительнотканных суставных сумках в большом количестве встречаются свободные нервные окончания, а также структуры, аналогичные тельцам Пачини и Руфини. Они чувствительны к растяжению и сжатию, возникающим при движении, и таким образом сигнализируют о положении тела в пространстве и движении отдельных его частей (кинестезия). Свободные нервные окончания могут, кроме того, воспринимать боль. Проводниковый и центральный отделы соматосенсорной системы. Нервные импульсы от рецепторов кожи и опорно-двигательною аппарата, кроме головы, по спинно-мозговым нервам достигают спинальных ганглиев, а затем через задние корешки поступают в спинной мозг. Афферентные волокна каждого заднего корешка проводят импульсы от определенной области тела — дерматома. Поступившая в спинной мозг информация используется в двух назначениях: она участвует в местных рефлексах, дуга которых замыкаются на уровне спинного мозга, а также передается в вышележащие отделы ЦНС по восходящим путям. При этом в восходящих трактах прослеживается соматотопическая организация: аксоны, присоединившиеся на более высоком уровне, располагаются со стороны серого вещества. Соответственно аксоны, идущие от нижней части тела, лежат более поверхностно.
Рис. 3.80. Соматосенсорные пути в спинном мозгу (по Melzack, 1965): / и 2 — миелинизированные и 2 — безмиелиновые волокна от механорецепторов (1 и 2) и болевых рецепторов (2); 3 — вставочный нейрон; 4 — проекционный нейрон; 5 — к переднему рогу (сгибательный рефлекс); 6 — к спиноталамическим трактам; /— V — пластины серого вещества Толстые миелиновые волокна огибают задний рог, отдают коллатерали к нейронам III—IV слоев и входят в задний канатик белого вещества. Как было установлено, большинство нейронов заднего рога получают афферентацию только одногс типа, однако существуют нейроны, на которых сходятся импульсы от различных рецепторов. На этом может быть основано взаимодействие различных рецепторных систем. Аксоны нейронов заднего рога могут уходить в белое вещество — в восходящие тракты, или достигать мотонейронов передних рогов и участвовать в осуществлении ряда спинальных рефлексов. Так импульсы от кожных рецепторов запускают сгибательный рефлекс. Он появляется при отдергивании конечности от болевого раздражителя (при ожоге и т. п.). Импульсы от рецепторов соматосенсорной системы проводятся по тонкому и клиновидному пучкам, а также по спинно-таламическим и спинно-мозжечковым путям и тройничной петле. Тонкий пучок несет импульсы от тела ниже V грудного сегмента) а клиновидный пучок — от верхней части туловища и рук. Эти пути образованы аксонами чувствительных нейронов, тела которых лежат в спинальных ганглиях, а дендриты образуют рецепторные окончания в коже, мышцах и сухожилиях. Пройдя весь спинной мозг и заднюю часть продолговатого, волокна тонкого и клиновидного пучков оканчиваются на нейронах тонкого и клиновидного ядер. Аксоны нейронов этих ядер идут по двум направлениям. Одни — под названием наружных дугообразных волокон — переходят на противоположную сторону, где в составе нижних ножек мозжечка оканчиваются на клетках коры червя. Нейриты последних связывают кору червя с ядрами мозжечка. Аксоны нейронов этих ядер, в составе нижних ножек мозжечка, направляются к преддверным ядрам моста. Другая, большая часть волокон от нейронов тонкого и клиновидного ядер спереди от центрального канала продолговатого мозга, совершает перекрест и образует медиальную петлю или лемниск. Поэтому оба этих пути называют лемнисковой системой. Медиальная петля идет через продолговатый мозг, покрышки моста и среднего мозга и заканчивается в латеральных и вентральных ядрах таламуса. По пути через ствол мозга волокна медиальной петли отдают коллатерали к ретикулярной формации. Волокна нейронов таламуса проходят в составе таламической лучистости к коре центральных областей больших полушарий. Как ядра продолговатого мозга, так и таламические и корковые проекции тонкого и клиновидного путей имеют соматотопическую организацию. По этим путям (особенно по клиновидному пучку) передается тонкая чувствительность от верхних конечностей, благодаря чему становятся возможными тонкие и точные движения пальцев руки. Этому способствует также наличие небольшого числа переключений с нейрона на нейрон — не происходит «растекания» возбуждения по структурам головного и спинного мозга. Спинно-таламический путь проводит возбуждение от рецепторов, раздражение которых вызывает болевые и температурные ощущения. Здесь также имеются волокна от суставных и тактильных рецепторов. Тела чувствительных нейронов этого пути также залегают в спинальных ганглиях. Центральные отростки этих нейронов входят в спинной мозг в составе задних корешков, где и оканчиваются на телах вставочных нейронов задних рогов на уровне IV— VI пластин. Аксоны нейронов задних рогов частично переходят на противоположную сторону, остальные остаются на своей стороне и в глубине бокового канатика образуют спинно-таламический путь. Последний проходит спинной мозг, покрышки продолговатого мозга, моста и ножек мозга и оканчивается на клетках вентрального ядра таламуса. По пути через ствол мозга от волокон этого тракта отходят коллатерали к ретикулярной формации. От таламуса волокна идут в составе таламической лучистости к коре, где оканчиваются, главным образом в постцентральной области. Спинно-мозжечковые задний и передний пути проводят возбуждение от проприорецепторов двигательного аппарата. Чувствительные нейроны этих путей расположены в спинальных ганглиях, а вставочные — в задних рогах спинного мозга. Нейриты вставочных нейронов, входящие в состав заднего спинно-мозжечкового пути, остаются на той же стороне спинного мозга в боковом канатике, а образующие передний путь переходят на противоположную сторону, где располагаются тоже в боковом канатике. Оба пути входят в мозжечок: задний — по его нижним ножкам, а передний — по верхним. Оканчиваются они на клетках коры червя. Отсюда импульсы идут по тем же путям, что и проходящие по наружным дугообразным волокнам из продолговатого мозга. Благодаря спинно-мозжечковым путям осуществляется интеграция информации от мышечных и суставных рецепторов конечностей и мозжечковых механизмов, необходимых для координации движений, поддержания мышечного тонуса и позы. Это особенно важно для работы нижних конечностей в положении стоя и при движении. Тройничная петля передает импульсы от механо-, термо- и болевых рецепторов головы. Чувствительными нейронами служат клетки тройничного узла. Периферические волокна этих клеток проходят в составе трех ветвей тройничного нерва, иннервирующих кожу лица (рис. 3.28). Центральные волокна чувствительных нейронов выходят из узла в составе чувствительного корешка тройничного нерва и проникают в мост в том месте, где он переходит в средние ножки мозжечка. В мосту эти волокна делятся Т-образно на восходящие и длинные нисходящие ветви (спинальный путь), которые оканчиваются на нейронах, образующих в покрышке моста основное сенсорное ядро тройничного нерва, а в продолговатом и спинном мозге — его спинальное ядро. Центральные волокна нейронов этих ядер совершают перекрест в верхней части моста и в качестве тройничной петли проходят по покрышке среднего мозга до таламуса, где оканчиваются самостоятельно или вместе с волокнами медиальной петли над клетками его вентрального ядра. Отростки нейронов этого ядра направляются в составе таламической лучистости к коре нижней части постцентральной области, Рис. 3.81. Соматосенсорная кора: 1 — нога;
3 — туловище; 4 — шея; 5 — голова; 6 — рука; 7 — локоть; 8 — предплечье; 9 — кисть; 10 — пальцы; 11 — большой палец; 12 — глаз; 13 — нос; 14 — лицо; 15 — губы; 16 — зубы; 17 — десны; 18 — челюсть; 19 — язык; 20 — глотка; 21 — внутренние органы
где главным образом и локализуется чувствительность, приходящая от структур головы. Соматосенсорные проекции в коре больших полушарий расположены в постцентральной извилине. Сюда подходят волокна от таламуса, приносящие импульсы от всех рецепторов кожи и опорно-двигательного аппарата. Здесь, также как и в таламусе, хорошо выражена соматотопическая организация проекций (рис. 3.81). Кроме первичной проекционной зоны, получающей афферентацию только от таламуса, существует и вторичная зона, на нейронах которой наряду с таламическими оканчиваются волокна от первичной зоны. В этой зоне происходит переработка сенсорных сигналов, отсюда они направляются в другие, в том числе и моторные области коры и подкорковые структуры.   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|


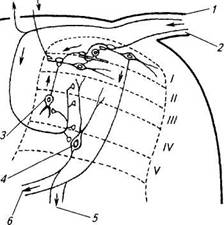 Как говорилось выше, серое вещество спинного мозга можно представить в виде пластин. Тонкие безмиелиновые волокна, подходящие к спинному мозгу от болевых и механорецепторов, оканчиваются в поверхностных пластинах, в основном в желатинозной субстанции. Тонкие миелиновые волокна доходят в основном только до краевой зоны (рис. 3.80).
Как говорилось выше, серое вещество спинного мозга можно представить в виде пластин. Тонкие безмиелиновые волокна, подходящие к спинному мозгу от болевых и механорецепторов, оканчиваются в поверхностных пластинах, в основном в желатинозной субстанции. Тонкие миелиновые волокна доходят в основном только до краевой зоны (рис. 3.80). 2 — бедро;
2 — бедро;