|
|
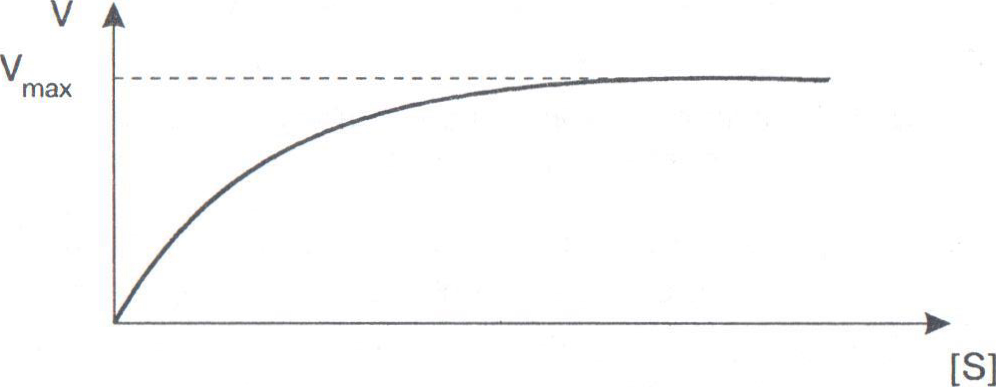
Биологические функции белков.Стр 1 из 11Следующая ⇒ Биологические функции белков. Структурная(строительная,пластическая). Эта функция заключается в том, что белки являются универсальным строительным материалом, из которого строятся все структурные образования организма, прежде всего все клетки и все внутриклеточные органоиды. Белки также входят в состав межклеточного вещества. Поэтому белков в организме много и на их долю в среднем приходится 1/6 часть от массы тела человека. Каталитическая. В организме имеются особые белки, выполняющие функции катализаторов химических реакций. Такие белки получили название ферменты, или энзимы. С помощью ферментов с большими скоростями протекают все химические реакции, составляющие обмен веществ. Сократительная. В основе всех форм движения и в первую очередь мышечного сокращения и расслабления лежит взаимодействие белков. Благодаря сократительной функции животные в отличие от растений могут произвольно перемещаться в пространстве. Регуляторная. Белки обладают амфотерностью и могут взаимодействовать как с кислотами, так и с основаниями. Поэтому белки являются важнейшими буферами организма, поддерживающими кислотность на необходимом уровне. Белки также участвуют в регуляции осмотического давления(гидростатическое давление, вызванное разностью концентраций. причиной возникновения осмотического давления является осмос- односторонняя диффузия молекул растворителя (чаще всего воды) через полупроницаемую мембрану в сторону раствора большей концентрации) и распределения воды между кровью и различными органами. Некоторые белки, являясь гормонами, непосредственно участвуют в регуляции обмена веществ. Рецепторная. В основе этой функции лежит способность белковых молекул реагировать на возникающие изменения условий внутренней среды организма. Например, имеются белки очень чувствительные к изменению концентраций различных веществ (хеморецепторы), к изменению температуры (терморецепторы), осмотического давления (осморецепторы), освещенности(фоторецепторы), положения мышцы в пространстве (проприорецепторы).Далее эта информация о возникающих изменениях поступает в центральную нервную систему. Кроме того, в органах, в которых проявляется действие гормонов, содержатся специальные белки- рецепторы гормонов. Эти белки позволяют данному органу избирательно извлекать из крови именно тот гормон, который в данных условиях необходим для регуляции обмена веществ в этом органе. Транспортная. Эта функция обусловлена тем, что белковые молекулы имеют большой размер, хорошо растворимы в воде и, перемещаясь по водным пространствам организма, могут переносить различные нерастворимые в воде соединения. Так, например, гемоглобин участвует в транспорте молекулярного кислорода от легких к различным органам, белки плазмы крови- альбумины- обеспечивают перенос жиров и жирных кислот. Защитная. Белки выполняют защитную функцию, участвуя в обеспечении иммунитета. К защитной функции часто относят участие белков в свертывании крови. В этом случае благодаря образованию тромба организм защищается от потери большого количества крови. Энергетическая. Окисление белков, как и всех других органических соединений, сопровождается выделением энергии. Однако роль белков как источников энергии невелика. В обычных условиях белки обеспечивают около 10% суточной потребности организма в энергии. Вклад белков в энергообеспечение организма возрастает при голодании и длительной физической работе, когда в организме исчерпываются запасы основных энергетических источников- углеводов и жиров. Зависимость скорости ферментативной реакции от концентрации фермента
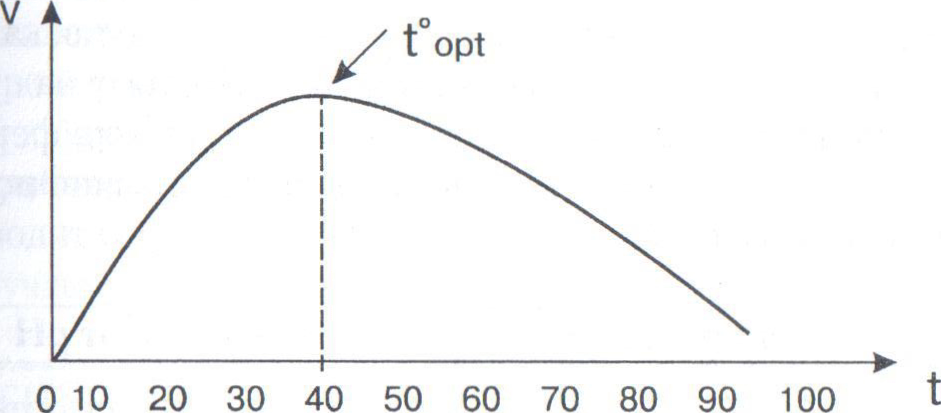
Зависимость скорости ферментативной реакции от температуры
Окисление жирных кислот. Жирные кислоты поступают с пищей или синтезируются в организме (кроме полиеновых кислот). Субстраты, необходимые для синтеза жирных кислот, образуются при катаболизме глюкозы и таким образом, часть глюкозы превращается сначала в жирные кислоты, а затем в жиры. Хотя специфический путь катаболизма жирных кислот заканчивается образованием ацетил-КоА, служащим исходным субстратом для синтеза жирных кислот, процессы синтеза и окисления жирных кислот необратимы. Они происходят в разных компартментах клеток (биосинтез протекает в цитозоле, а окисление - в митохондриях) и катализируются разными ферментами. Окисление жирных кислот как источников энергии увеличивается в постабсорбтивный период, при голодании и физической работе. В этих состояниях их концентрация в крови увеличивается в результате мобилизации из жировых депо, и они активно окисляются печенью, мышцами и другими тканями. При голодании часть жирных кислот в печени превращается в другие "топливные" молекулы - кетоновые тела. Они, в отличие от жирных кислот, могут использоваться нервной тканью как источник энергии. При голодании и длительной физической работе кетоновые тела служат источником энергии для мышц и некоторых других тканей. β-Окисление жирных кислот β-Окисление - специфический путь катаболизма жирных кислот, при котором от карбоксильного конца жирной кислоты последовательно отделяется по 2 атома углерода в виде ацетил-КоА. Метаболический путь - β-окисление - назван так потому, что реакции окисления жирной кислоты происходят у β-углеродного атома. Реакции β-окисления и последующего окисления ацетил-КоА в ЦТК служат одним из основных источников энергии для синтеза АТФ по механизму окислительного фосфорилирования. β-Окисление жирных кислот происходит только в аэробных условиях.
37.Кетоновые тела - это способ транспорта ацетильной группы При состояниях, сопровождающихся снижением глюкозы крови, клетки органов и тканей испытывают энергетический голод. Так как окисление жирных кислот процесс "трудоемкий", анервная ткань вообще неспособна окислять жирные кислоты, то печень облегчает использование этих кислот тканями, заранее окисляя их до уксусной кислоты и переводя последнюю в транспортную форму – кетоновые тела. К кетоновым телам относят три соединения близкой структуры – ацетоацетат, 3-гидроксибутират и ацетон. тимулом для образования кетоновых тел служит поступление большого количества жирных кислот в печень. Как уже указывалось, при состояниях, активирующих липолиз в жировой ткани, не менее 30% образованных жирных кислот задерживаются печенью. К таким состояниям относится голодание, сахарный диабет I типа, длительные физические нагрузки. Так как синтез ТАГ в этих условиях невозможен, то жирные кислоты из цитозоля попадают в митохондрии и окисляются с образованием кетонов. Кроме отмеченных ситуаций, количество кетоновых тел в крови возрастает при алкогольном отравлении и потреблении жирной пищи. При богатой жирами диете, особенно у детей, жирные кислоты не успевают включиться в состав ТАГ и ЛПОНП и частично переходят в митохондрии, что увеличивает синтез кетоновых тел. Приалкогольном отравлении субстратом для синтеза кетонов является ацетил-SКоА, синтезируемый при обезвреживании этанола. В обычных условиях синтез кетоновых тел также идет, хотя в гораздо меньшем количестве. Для этого используются как жирные кислоты, так и безазотистые остатки кетогенных и смешанныхаминокислот. Синтез кетоновых тел (кетогенез) Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо спонтанно декарбоксилируется до ацетона. Далее все три соединения поступают в кровь и разносятся по тканям. Ацетон, как летучее вещество, легко удаляется с выдыхаемым воздухом и потом. Все кетоновые тела могут выделяться с мочой. Используются кетоновые тела клетками всех тканей, кроме печени и эритроцитов. Особенно активно, даже в норме, они потребляются миокардом и корковым слоем надпочечников. Реакции утилизации кетоновых тел примерно совпадают с обратным направлением реакций синтеза. В цитозоле 3-гидроксибутират окисляется, образующийся ацетоацетат проникает в митохондрии, активируется за счет сукцинил-SКоА и превращается в ацетил-SКоА, который сгорает в ЦТК.
38.Синтез жирных кислот и жира Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме пече-ночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-кар-боксилаза. Фермент содержит в качестве простетической группы биотин. Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА. Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты. Последовательность реакций, происходящих при синтезе жирных кислот Далее цикл реакций повторяется. По сравнению с β-окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитозоле клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путем связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ); при биосинтезе образуется D(–)-изомер 3-гидроксикис-лоты, а не L(+)-изомер, как это имеет место при β-окислении жирных кислот; необходимость для синтеза жирных кислот кофермента НАДФН. . Холестери́н -холестерол — органическое соединение, природ-ный жирный (липофильный) спирт, содержащийся в клеточных мембранах всех животных организмов за исключением безъядер-ных (прокариот). Нерастворим в воде, растворим в жирах и органических растворителях. Биологическая роль. Холестерин в составе клеточной плазматической мембраны играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов. Таким образом, холестерин — стабилизатор текучести плаз-матической мембраны. Холестерин открывает цепь биосинтеза стероидных половых гормонов и кортикостероидов, служит основой для образования жёлчных кислот и витаминов группы D, участвует в регулировании проницаемости клеток и предохраняет эритроциты крови от действия гемолитических ядов. Обмен холестерина. Свободный холестерин подвергается окислению в печени и органах, синтезирующих стероидные гормоны (надпочечники, семенники, яичники, плацента). Это единственный процесс необратимого выведения холестерина из мембран и липопротеидных комплексов. Ежедневно на синтез стероидных гормонов расходуется 2—4% холестерина. В гепатоцитах 60—80% холестерина окисляется до желчных кислот, которые в составе желчи выделяются в просвет тонкой кишки и участвуют в пищеварении (эмульгировании жиров). Вместе с желчными кислотами в тонкую кишку выделяется небольшое количество свободного холестерина, который частично удаляется с каловыми массами, а оставшаяся часть его растворяется и вместе с желчными кислотами и фосфолипидами всасывается стенками тонкой кишки. Желчные кислоты обеспечивают разложение жиров на составные части (эмульгиро-вание жиров). После выполнения этой функции 70—80% остав-шихся желчных кислот всасывается в конечном отделе тонкой кишки (подвздошной кишке) и поступает по системе воротной вены в печень. Здесь стоит отметить, что желчные кислоты имеют еще одну функцию: они являются важнейшим стимулятором поддержания нормальной работы (моторики) кишечника. В печени начинают синтезироваться не до конца сформировавшиеся (насцентные) липопротеиды высокой плотности. Окончательно ЛПВП формируются в крови из специ-альных белков (апобелков) хиломикронов, ЛПОНП и холесте-рина, поступающего из тканей, в том числе и из артериальной стенки. Более просто кругооборот холестерина можно объяснить следующим образом: холестерин в составе липопротеидов несет жир из печени к различным частям вашего тела, используя кровеносные сосуды в качестве транспортной системы. После доставки жира холестерин возвращается в печень и повторяет свою работу снова. Первичные желчные кислоты. (холевая и хенодезоксихолевая) синтезируются в гепатоцитах печени из холестерина. Вторичная: дезоксихолевая кислота (первоначально синтезируется в толстой кишке). Желч-ные кислоты образуются в митохондриях гепатоцитов и вне их из холестерина с участием АТФ. Гидроксилирование при образова-нии кислот осуществляется в эндоплазматическом ретикулуме гепатоцита. Первичный синтез желчных кислот ингибируется (тормозится) желчными кислотами, присутствующими в крови. Однако, если всасывание в кровь желчных кислот будет недоста-точно, например, из-за тяжёлого поражения кишечника, то печень, способная произвести не более 5 г желчных кислот в сутки, не сможет восполнить требуемое для организма количество желчных кислот. Желчные кислоты — главные участ-ники энтерогепатической циркуляции у человека. Вторичные желчные кислоты (дезоксихолевая, литохолевая, урсодезоксихолевая, аллохолевая и другие) образуются из первичных желчных кислот в толстой кишке под действием кишечной микрофлоры. Их количество невелико. Дезоксихолевая кислота всасывается в кровь и секретируется печенью в составе желчи. Литохолевая кислота всасывается значительно хуже, чем дезоксихолевая
39. Строение мононуклеотидов. По своему строению нуклеиновые кислоты являются полинуклеотидами, состоящими из мононуклеотидов или нуклеотидов. Нуклеотид сложное органическое соединение, состоящее из трех частей: азотистого основания, углевода и остатков фосфорной кислоты. Азотистые основания - это гетероциклические органические соединения, относящиеся к двум классам – пурины и пиримидины. Из пуринов в состав нуклеиновых кислот входят аденин и гуанин А из пиримидинов цитозин, тимин (ДНК) и урацил (РНК) Углеводом, входящим в состав нуклеотидов может быть рибоза (РНК) и дезоксирибоза (ДНК) Азотистое основание, связанное с углеводом называется нуклеозидом. Фосфорная кислота присоединяется эфирной связью к пятому атому углерода рибозы или дезоксирибозы. Нуклеотиды, входящие в состав нуклеиновых кислот имеют один остаток фосфорной кислоты и называются мононуклеотидами. Однако в клетке встречаются ди- и тринуклеотиды. Биологическая роль нуклеиновых кислот и функции мононуклеотидов. 1. ДНК: хранение генетической информации. 2. РНК: - хранение генетической информации (информосомы, некоторые РНК-вирусы); - реализация генетической информации: и-РНК (м-РНК) — информационная (матричная), т-РНК (транспортная), р-РНК (рибосомальная). Участвуют в процессе синтеза белка; - каталитическая функция: некоторые молекулы РНК катализируют реакции гидролиза 3′,5′-фосфодиэфирной связи в самой молекуле РНК-«самосплайсинг». Функции мононуклеотидов: 1) структурная — построение нуклеиновых кислот, некоторых коферментов и простетических групп ферментов; 2) энергетическая — аккумуляторы энергии за счет имеющихся макроэргических связей. АТ Ф — универсальный аккумулятор энергии, энергия УТ Ф используется для синтеза гликогена, ЦТ Ф — для синтеза липидов, ГТФ — для движения рибосом в ходе трансляции (биосинтез белка) и передачи гормонального сигнала (G-белок); 3) регуляторная: мононуклеотиды — аллостерические эффекторы многих ключевых ферментов, цАМФ и цГМФ — посредники в передаче гормонального сигнала при действии многих гормонов на клетку (аденилатциклазная система), активируют протеинкиназы.
Внутриклеточный протеолиз Белки, входящие в состав клеток организма, также подвергаются постоянному распаду под влиянием внутриклеточных протеолитических, называемых внутриклеточными протеиназами, или катепсинами. Эти ферменты локализованы в специальных внутриклеточных органоидах – лизосомах. Мембраны, окружающие лизосомы непроницаемы для катепсинов, зато пропускают внутрь лизосом белки, содержащие протеолизу. По своему действию катепсины похожи на ферменты желудочного и поджелудочного соков: катепсины вызывают превращение белков в олигопептиды, которые легко выходят из лизосом в цитоплазму клеток и превращаются там в аминоксилоты. Такой распад протекает с участием цитоплазматических ферментов, аналогичных ферментам желудочного сока. В цитоплазме клеток организма были обнаружены особые белковые мультиферментные комплексы, названные протеосомами. Протеосомы предназначены для избирательного расщепления белков, которых не должно быть в клетках. Такому превращению подвергаются чужеродные и дефектные ьелки, а также регуляторные белки, присутствие которых ограничено во времени (например, белки-гормоны, белки-регуляторы транскрипции). Для предупреждения чрезмерного распада собственных белковв организме имеются особые белки – эндогенные ингибиторы протеиназ, снижающие скорость протеолиза. Особенно много таких ингибиторов протеолиза в богатой белками плазме крови. В сутки внутриклеточному протеолизу подвергается 200-300 г. собственных белков организма, что приводит к возникновению примерно такого же количества аминокислот. Важно отметить, что при распаде как пищевых, так и собственных белков организма образуются аминокислоты одних и тех же 20 видов. Поэтому в течение суток в организме появляется около 300-400 г свободных аминокислот. Глубокий распад белков, сопровождающийся расщеплением всех пептидных связей и приводящий к образованию аминокислот, называется тотальным протеолизом. В некоторых случаях под воздействием протеолитических ферментов в молекуле белка избирательно расщепляется лишь одна пептидная связь между строго определенными аминокислотами и от белка отщепляется часть его молекулы – полипетид. Такой распад белка называется ограниченным протеолизом. Примером ограниченного протеолиза является превращение проферментов в ферменты (например, активация протеолитических ферментов, участвующих в пищеварении; переход факторов свертывания крови из неактивной формы в активную.
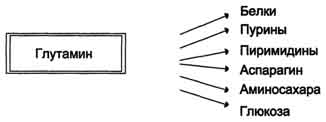
Синтез белка Большая часть аминокислот используется для синтеза белков. В организме взрослого человека существует равновесие между распадом и синтезом белков. У детей преобладает синтез, ведущий к накоплению белков в организме, что является обязательным условием роста и развития организма. Синтез белков происходит при обязательном участии нуклеиновых кислот. 1 этап синтеза белка - транскрипция - осуществляется в клеточном ядре с использованием ДНК как источника генетической информации. Генетическая (наследственная) информация обусловливает порядок расположения аминокислот в полипептидных цепях синтезируемого белка. Эта информация закодирована строгой последовательностью азотистых оснований мононуклеотидов в молекуле ДНК. Каждая аминокислота кодируется сочетанием трех азотистых оснований, называемым кодоном, или триплетом. Участок молекулы ДНК, содержащий информацию об определенном белке, получил название «ген». На этом участке ДНК во время транскрипции по принципу комплементарности синтезируется информационная РНК (иРНК). Эта нуклеиновая кислота представляет собой копию соответствующего гена. Следовательно, иРНК содержит информацию о строении белка, закодированного в данном гене. Образовавшаяся иРНК выходит из ядра и поступает в цитоплазму. Аналогичным образом на ДНК как на матрице происходит синтез рибосомных (рРНК) и транспортных (тРНК) РНК. 2 этап — рекогниция (распознование), протекающего в цитоплазме, аминокислоты избирательно связываются со своими переносчиками - транспортными РНК (тРНК). Все тРНК (их обнаружено около 60) построены сходным образом. Молекула каждой тРНК представляет собой короткую полинуклеотидную цепь, содержащую примерно 80 нуклеотидов и частично закрученную в двойную спираль, что приводит к возникновению пространственной конфигурации «изогнутого клеверного листа». На одном конце полинуклеотидной цепи у всех тРНК находится нуклеотид, содержащий аденин. К этому концу молекулы тРНК присоединяется аминокислота. Петля, противоположная месту присоединения аминокислоты, содержит антикодон, состоящий из трех азотистых оснований и предназначенный для последующего связывания с комплементарным кодоном иРНК. Одна из боковых петель молекулы тРНК обеспечивает присоединение тРНК к ферменту, участвующему в рекогни- ВДи> а другая, боковая, петля необходима для присоединения тРНК к рибосоме на следующем этапе синтеза белка. На этом этапе в качестве источника энергии используется молекула АТф. в результате рекогниции образуется комплекс аминокислота-тРНК (аминоацил-тРНК): аминокислота + тРНК ----------˃ аминоацил-тРНК В составе этого комплекса аминокислота обладает повышенной химической активностью. В связи с этим второй этап синтеза белка часто называют активацией аминокислот. 3 этап синтеза белка - трансляция - происходит на рибосомах. Каждая рибосома состоит из двух частей - большой и малой субчастиц. По химическому составу обе субчастицы представляют собою нук- леопротеиды, состоящие из рибосомных РНК и белков. Рибосомы способны легко распадаться на субчастицы (диссоциация), которые снова могут соединяться друг с другом, образуя рибосому (ассоциация). Трансляция начинается с диссоциации рибосомы на субчастицы, которые сразу же присоединяются к начальной части молекулы информационной РНК, поступающей из ядра. При этом между субчастицами остается пространство (так называемый туннель), где располагается небольшой участок иРНК. Затем к образовавшемуся комплексу рибосома - иРНК присоединяются тРНК, связанные с аминокислотами. Присоединение тРНК к этому комплексу происходит путем связывания одной из боковых петлей тРНК с рибосомой и связывания антикодона тРНК с комплементарным ему кодоном иРНК, находящимся в туннеле между субчастицами рибосомы. Одновременно к комплексу рибосома - иРНК могут присоединиться только две тРНК с аминокислотами. Благодаря специфическому связыванию антикодонов тРНК с ко донами иРНК, к участку молекулы иРНК, находящемуся в туннеле, присоединяются только молекулы тех тРНК, у которых антикодоны комплементарны кодонам иРНК. Поэтому эти тРНК доставляют в рибосомы только строго определенные аминокислоты. Далее аминокислоты соединяются друг с другом пептидной связью и образуется дипептид, который связан с одной из тРНК. После этого рибосома передвигается вдоль иРНК ровно на один кодон (это перемещение рибосомы называется транслокацией). В результате транслокации свободная (без аминокислоты) тРНК отщепляется от рибосомы, а в зоне туннеля появляется новый кодон, к которому присоединяется по принципу комплементарности еще одна тРНК с аминокислотой, соответствующей этому кодону. Доставленная аминокислота соединяется с ранее образовавшимся дипептидом, что приводит к удлинению пептидной цепи. Далее следуют новые транслокации, поступление на рибосому новых тРНК с аминокислотами и дальнейшее удлинение пептидной цепи. Таким образом порядок включения аминокислот в синтезируемый белок определяется последовательностью кодонов в иРНК. Завершается синтез полипептидной цепи при поступлении в туннель особого кодона, который не кодирует аминокислоты и к которому не может присоединиться ни одна тРНК. Такие кодоны называются терминирующими, или нонсенс-кодонами. Особенно велика их роль в синтезе белков, молекула которых состоит из нескольких полипептидов. В итоге за счет описанных трех этапов синтезируются полипептиды, т. е. формируется первичная структура белка. Высшие (пространственные) структуры (вторичная, третичная, четвертичная) возникают самопроизвольно. Это объясняется тем, что пространственные структуры фиксируются в основном химическими связями, возникающими между радикалами аминокислот. Поэтому в каждом белке эти связи образуются специфически в соответствии с генетически обусловленной последовательностью расположения радикалов в полипептидных цепях. В некоторых случаях полипептидная цепь после завершения синтеза подвергается незначительной химической модификации, в результате чего в ней появляются некодируемые аминокислоты (не относящиеся к 20 обычным аминокислотам). Например, при синтезе белка коллагена на рибосомах образуется его предшественник - проколлаген, содержащий в большом количестве аминокислоты лизин и пролин. Эти аминокислоты, находящиеся в составе полипептидной цепи, подвергаются в ходе модификации окислению и превращаются соответственно в окси- лизин и оксипролин, что приводит к переходу проколлагена в коллаген. Синтез белков - процесс энергоемкий. Для включения в молекулу синтезируемого белка только одной аминокислоты требуется не менее трех молекул АТФ. В очень небольших количествах белки еще синтезируются в митохондриях, где также имеются ДНК, иРНК, тРНК и рибосомы. Синтез белков в организме ускоряется соматотропным гормоном (гормоном роста) и тестостероном. Тормозится синтез белков гормонами коры надпочечников - глюкокортикоидами. Регулирующее действие всех этих гормонов связано с их влиянием на скорость транскрипции. Синтез белков подавляют многие антибиотики, ингибирующие трансляцию. Рис. 9-10. Метаболизм азота глутамина в кишечнике. В почках также происходит гидролиз глутамина под действием глутаминазы с образованием аммиака. Этот процесс является одним из механизмов регуляции кислотно щелочного равновесия в организме и сохранения важнейших катионов для поддержания осмотического давления. Глутаминаза почек значительно индуцируется при ацидозе, образующийся аммиак нейтрализует кислые продукты обмена и в виде аммонийных солей экскретируется с мочой (рис. 9-11). Эта реакция защищает организм от излишней потери ионов Na+ и К+, которые также могут использоваться для выведения анионов и утрачиваться. При алкалозе количество глутаминазы в почках снижается. В почках образуется и выводится около 0,5 г солей аммония в сутки. Высокий уровень глутамина в крови и лёгкость его поступления в клетки обусловливают использование глутамина во многих анаболических процессах. Глутамин - основной донор азота в организме. Амидный азот глутамина используется для синтеза пуриновых и пиримидиновых
Рис. 9-11. Метаболизм амидного азота глутамина в почках. нуклеотидов, аспарагина, аминосахаров и других соединений (рис. 9-12).
Рис. 9-12. Пути использования глутамина в организме. Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
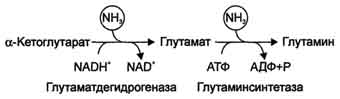
Существуют 2 изоформы этого фермента - глутаминзависимая и аммиакзависимая, которые используют разные доноры амидных групп. Первая функционирует в животных клетках, вторая преобладает в бактериальных клетках, но присутствует и у животных. Однако такой путь обезвреживания аммиака в клетках человека используется редко и к тому же требует больших энергетических затрат (энергию двух макроэргических связей), чем синтез глутамина. Наиболее значительные количества аммиака обезвреживаются в печени путём синтеза мочевины. В первой реакции процесса аммиак связывается с диоксидом углерода с образованием карбамоилфосфата, при этом затрачиваются 2 молекулы АТФ. Реакция происходит в митохондриях гепатоцитов под действием фермента карбамоилфос-фатсинтетазы I. Карбамоилфосфатсинтетаза II локализована в цитозоле клеток всех тканей и участвует в синтезе гшримидиновых нуклеотидов (см. раздел 10). Карбамоилфосфат затем включается в орнитиновый цикл и используется для синтеза мочевины. В мозге и некоторых других органах может протекать восстановительное аминирование α -кетоглутарата под действием глутаматдегидрогеназы, катализирующей обратимую реакцию. Однако этот путь обезвреживания аммиака в тканях используется слабо, так как глутаматдегидрогеназа катализирует преимущественно реакцию дезаминирования глутамата. Хотя, если учитывать последующее образование глутамина, реакция выгодна для клеток, так как способствует связыванию сразу 2 молекул NH3.
Из мышц и кишечника избыток аммиака выводится преимущественно в виде аланина. Этот механизм необходим, так как активность глутаматдегидрогеназы в мышцах невелика и непрямое дезаминирование аминокислот малоэффективно. Поэтому в мышцах существует ещё один путь выведения азота. Образование аланина в этих органах можно представить следующей схемой (см. схему ниже). Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, основным источником которого служит процесс окисления глюкозы. Мышцы выделяют особенно много аланина в силу их большой массы, активного потребления
Схема глюкозы при физической работе, а также потому, что часть энергии они получают за счёт распада аминокислот. Образовавшийся аланин поступает в печень, где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается, а пируват включается в глюконеогенез. Глюкоза из печени поступает в ткани и там, в процессе гликолиза, опять окисляется до пирувата (рис. 9-13). Образование аланина в мышцах, его перенос в печень и перенос глюкозы, синтезированной в печени, обратно в мышцы составляют глюкозо-аланиновый цикл, работа которого сопряжена с работой глюкозо-лактатного цикла (см. раздел 7). Совокупность основных процессов обмена аммиака в организме представлена на рис. 9-14. Доминирующими ферментами в обмене аммиака служат глутаматдегидрогеиаза и глутаминсинтетаза.
Вопрос 50. Биологическая роль витаминов. Основные причины гиповитаминозов.Биологическая роль- они входят в состав коферментов и простетических групп ферментов, и следовательно используются организмом как строительный материал при синтезе соответствующих небелковых частей ферментов.Гиповитаминоз- специфическое заболевание протекающие в более легкой форме по сравнению в авитаминозами, вызываемым недостаточным содержанием отдельных витаминов в организме.Причины: Экзогенные(связанные с питанием)неправильное приготовление пищи, приготовление пищи с малым кол-вом витаминов, однообразное питание. Эндогенные (связанные с состоянием организма)заболевание ЖКТ и печени, угнетение микрофлоры кишечника, повышенная потребность в витаминах(напр: беременность)
50. Биологическая роль витаминов, основные причины гиповитаминоза. Биологическая роль витаминов. Витамины – это органические соединения с низкомолекулярной структурой. Поступают в организм, в основном, с пищей, так как организм их синтезирует в крайне ограниченных количествах. Виды витаминов: · Водорастворимые витамины (витамины группы В: В1, В2 ,В6, В12, ВС; С; РР; Р; Н). Эти витамины участвуют в образовании различных коферментов. · Жирорастворимые витамины (A1, D2, D3,К и Е) участвуют в определении и поддержании функциональности субклеточных структур и клеточных мембран. При значительном дефиците витаминов все процессы в организме не могут протекать в нормальном режиме, что вызывает нарушения в деятельности органов и их систем. Витамин А (ретинол) нужен для поддержания красивой кожи, волос и всех слизистых, нормальной работы зрительной системы. Без него невозможно гармоничное формирование организма в период отрочества. · Витамин В1 (тиамин) координирует углеводный обмен, поставляющий в организм энергию, поддерживает работу нервной, пищеварительной, дыхательной системы. · Витамин В2 (рибофлавин) отвечает за способность клеток к восстановлению, поэтому при его недостатке даже маленькие кожные трещинки заживают с трудом. Незаменима его функция в процессах окисления и синтеза в организме, а также в поддержании функциональности вегетативного отдела нервной системы. · Витамин В6 (пиридоксин) – участник обмена белков и жиров, стимулирующий использование организмом природных антиоксидантов в виде ненасыщенных жирных кислот. Определенная доля этого витамина образуется микрофлорой кишечника. · Витамин В12 (цианокобаламин) принимает важное участие в процессах кроветворения и белковом обмене. Благодаря этому витамину каротин усваивается организмом, переходя в витамин А. Образуется в толстом кишечнике. · Витамины группы D участвуют в кальциево-фосфорном обмене, поддержке здоровья эндокринных желез. При недостатке - происходит нарушение образования зубов и костей, поражаются мышцы, ухудшается работа пищеварительной, ССС и НС. · Витамин С является важным компонентом окислительно-восстановительных процессов, препятствующим образованию опухолей. Без него не обходятся процессы кроветворения, усвоения железа. Он нужен для поддержки иммунитета. · Витамин Е (токоферола ацетат) – природный антиоксидант, поддерживающий репродуктивные функции. · Витамин РР – один из основных регуляторов обмена веществ, при недостатке которого большинство тканей и органов подвергаются патологическим изменениям. Причины гиповитаминова. · Нехватка витамина в рационе, несбалансированное питание · Разрушение питательных веществ в содержащей их пище вследствие нарушений условий хранения или в результате температурной или иной кулинарной обработки · Действие веществ-антагонистов, которые содержатся в тех или иных продуктах и приводят к разрушению витаминов, нарушению их усвоения (в частности, белок яйца затрудняет усвоение биотина). Гиповитаминоз также может быть обусловлен эндогенными (внутренними причинами): - расстройства работы желудочно-кишечного тракта, приводящие к нарушению процесса всасывания и усвоения витаминов. Часто недостаточное усвоение витаминов бывает вызвано дисбактериозом — нарушением микрофлоры кишечника, которое часто сопровождает длительный прием антибиотиков, прохождение химиотерапии при туберкулезе, онкологических болезнях. Микрофлора участвует в процессе синтеза некоторых витаминов, который происходит внутри организма; ухудшение ее состояния, наличие в ней патогенных микроорганизмов, а также глистные и другие паразитарные инвазии приводят к резкому сокращению запасов витамина в организме. - генетически обусловленные дефекты ферментных систем, транспортных функций, обеспечивающих всасывание и распределение витаминов. - прием некоторых лекарственных препаратов также может вызвать гиповитаминоз. - увеличение потребности человека в витаминах(беременность и кормление, периоды повышенной физич   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|