|
|
Общие пути распада аминокислотЧасть аминокислот подвергается распаду и превращается в конечные продукты: С02, Н20 и NH3. Распад начинается с реакций, общих для большинства аминокислот. К ним относятся: а) декарбоксилирование - отщепление от аминокислот карбоксильной группы в виде углекислого газа:
Это превращение аминокислот обычно протекает с очень низкой скоростью и аминов образуется мало. Но некоторые амины, находясь в очень низкой концентрации, обладают высокой биологической активностью и влияют на различные функции организма. Примером такого амина является гистамин, образующийся из аминокислоты гистидина. б) дезаминирование - отщепление аминогруппы в виде NH3. У человека дезаминирование аминокислот идет окислительным путем:
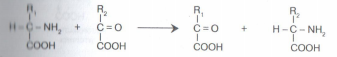
Дезаминирование аминокислот также протекает с низкой скоростью. И только одна аминокислота – глутаминовая – дезаминируется с высокой скоростьювследствие наличияв организме активного фермента, вызывающего дезаминирование только этой аминокислоты. в) трансаминирование (переамииирование) - реакция между аминокислотами и а-кетокислотами. В ходе этой реакции ее участники обмениваются функциональными группами, в результате чего аминокислота превращается в а-кетокислоту, а кетокислота становится аминокислотой:
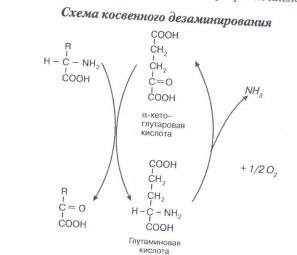
Трансаминированию подвергаются все аминокислоты. В этой реакции участвует кофермент - фосфопиридоксаль, для образования которого необходим витамин В6 - пиридоксин. Трансаминирование - это главное превращение аминокислот в организме, так как его скорость значительно выше, чем у реакций декарбоксилирования и дезаминирования. Трансаминирование выполняет две основные функции: а) за счет трансаминирования одни аминокислоты могут превращаться в другие. При этом общее количество аминокислот не меняется, но изменяется соотношение между ними. С пищей в организм поступают чужеродные белки, у которых аминокислоты находятся в иных пропорциях по сравнению с белками организма. Путем трансаминирования происходит корректировка аминокислотного состава организма. б) является составной частью косвенного (непрямого) дезаминирования аминокислот - процесса, с которого начинается распад большинства аминокислот. На первой стадии этого процесса аминокислоты вступают в реакцию трансаминирования с а-кетоглутаровой кислотой (а-кетокислота). Аминокислоты при этом превращаются в а- кетокислоты, а а-кетоглутаровая кислота переходит в глутаминовую кислоту (аминокислота). На второй стадии появившаяся глутаминовая кислота подвергается дезаминированию, от нее отщепляется NH3 и снова образуется а-кетоглутаровая кислота.
Итоговое уравнение косвенного дезаминирования совпадает с уравнением прямого дезаминирования. Однако у косвенного дезаминирования скорость значительно выше прямого, что обусловлено высокой активностью ферментов, катализирующих обе стадии этого процесса. Отсюда следует, что реакцией, с которой начинается распад аминокислот, является трансаминирование. Образовавшиеся а-кетокислоты далее подвергаются глубокому распаду и превращаются в конечные продукты С02 и Н20. Для каждой из 20 кетокислот (их образуется столько же, сколько имеется видов аминокислот) имеются свои специфические пути распада. Однако при распаде некоторых аминокислот в качестве промежуточного продукта образуется пировиноградная кислота, из которой возможен синтез глюкозы. Поэтому аминокислоты, из которых возникают такие кетокислоты, получили название глюкогенные. Другие же кетокислоты при своем распаде не образуют пирувата. Промежуточным продуктом у них является ацетилкофермент А, из которого невозможно получить глюкозу, но зато могут синтезироваться кетоновые тела. Аминокислоты, соответствующие таким кетокислотам, называются кетогенные. Второй продукт косвенного дезаминирования аминокислот - аммиак. Для организма аммиак является высокотоксичным. Поэтому в организме имеются молекулярные механизмы его обезвреживания.
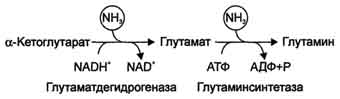
Вопрос49. Обезвреживание аммиака. синтез глутаминовой кислоты (восстановительное аминирование) – взаимодействие α-кетоглутарата с аммиаком. Реакция по сути обратна реакции окислительного дезаминирования, однако в качестве кофермента используется НАДФН. Происходит практически во всех тканях, кроме мышечной, но имеет небольшое значение, т.к. для глутаматдегидрогеназы предпочтительным субстратом является глутаминовая кислота и равновесие реакции сдвинуто в сторону α-кетоглутарата,
Реакция синтеза глутаминовой кислоты синтез глутамина – взаимодействие глутамата с аммиаком. Является главным способом уборки аммиака, наиболее активно происходит в нервной и мышечной тканях, в почках, сетчатке глаза, печени. Реакция протекает в митохондриях.
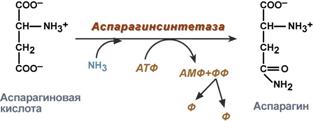
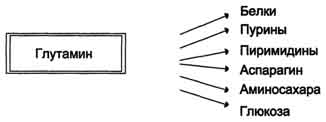
Реакция синтеза глутамина Образование большого количества глутамина обеспечивает высокие концентрации его в крови (0,5-0,7 ммоль/л). Так как глутамин проникает через клеточные мембраны путем облегченной диффузии, то он легко попадает не только в гепатоциты, но и в другие клетки, где есть потребность в аминогруппах. Азот, переносимый глутамином, используется клетками для синтеза пуринового и пиримидинового колец, гуанозинмонофосфата (ГМФ), аспарагина, глюкозамино-6-фосфата (предшественник всех остальных аминосахаров). синтез аспарагина – взаимодействие аспартата с аммиаком. Является второстепенным способом уборки аммиака, энергетически невыгоден, т.к. при этом тратятся 2 макроэргические связи,
Реакция синтеза аспарагина синтез карбамоилфосфата в митохондриях печени – реакция является первой в процессе синтеза мочевины, средства для удаления аммиака из организма.
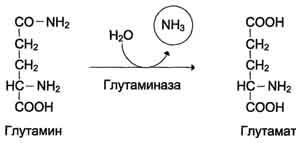
Вопрос: 49 Обезвреживание аммиака. Высокая интенсивность процессов дезаминирования аминокислот в тканях и очень низкий уровень аммиака в крови свидетельствуют о том, что в клетках активно происходит связывание аммиака с образованием нетоксичных соединений, которые выводятся из организма с мочой. Эти реакции можно считать реакциями обезвреживания аммиака. В разных тканях и органах обнаружено несколько типов таких реакций. Основной реакцией связывания аммиака, протекающей во всех тканях организма, является синтез глутамина под действием глутамин-синтетазы:
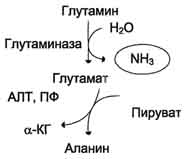
Глутаминсинтетаза локализована в митохондриях клеток, для работы фермента необходим кофактор - ионы Mg2+. Глутаминсинтетаза - один из основных регуляторных ферментов обмена аминокислот и аллостерически ингибируется АМФ, глюкозо-6-фосфатом, а также Гли, Ала и Гис. Глутамин легко транспортируется через клеточные мембраны путём облегчённой диффузии (для глутамата возможен только активный транспорт) и поступает из тканей в кровь. Основными тканями-поставщикам:и глутамина служат мышцы, мозг и печень. С током крови глутамин транспортируется в кишечник и почки. В клетках кишечника под действием фермента глутаминазы происходит гидролитическое освобождение амидного азота в виде аммиака:
Образовавшийся в реакции глутамат подвергается трансаминированию с пируватом. ос-Аминогруппа глутаминовой кислоты переносится в состав аланина (рис. 9-10). Большие количества аланина поступают из кишечника в кровь воротной вены и поглощаются печенью. Около 5% образовавшегося аммиака удаляется в составе фекалий, небольшая часть через воротную вену попадает в печень, остальные ~90% выводятся почками.
Рис. 9-10. Метаболизм азота глутамина в кишечнике. В почках также происходит гидролиз глутамина под действием глутаминазы с образованием аммиака. Этот процесс является одним из механизмов регуляции кислотно щелочного равновесия в организме и сохранения важнейших катионов для поддержания осмотического давления. Глутаминаза почек значительно индуцируется при ацидозе, образующийся аммиак нейтрализует кислые продукты обмена и в виде аммонийных солей экскретируется с мочой (рис. 9-11). Эта реакция защищает организм от излишней потери ионов Na+ и К+, которые также могут использоваться для выведения анионов и утрачиваться. При алкалозе количество глутаминазы в почках снижается. В почках образуется и выводится около 0,5 г солей аммония в сутки. Высокий уровень глутамина в крови и лёгкость его поступления в клетки обусловливают использование глутамина во многих анаболических процессах. Глутамин - основной донор азота в организме. Амидный азот глутамина используется для синтеза пуриновых и пиримидиновых
Рис. 9-11. Метаболизм амидного азота глутамина в почках. нуклеотидов, аспарагина, аминосахаров и других соединений (рис. 9-12).
Рис. 9-12. Пути использования глутамина в организме. Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
Существуют 2 изоформы этого фермента - глутаминзависимая и аммиакзависимая, которые используют разные доноры амидных групп. Первая функционирует в животных клетках, вторая преобладает в бактериальных клетках, но присутствует и у животных. Однако такой путь обезвреживания аммиака в клетках человека используется редко и к тому же требует больших энергетических затрат (энергию двух макроэргических связей), чем синтез глутамина. Наиболее значительные количества аммиака обезвреживаются в печени путём синтеза мочевины. В первой реакции процесса аммиак связывается с диоксидом углерода с образованием карбамоилфосфата, при этом затрачиваются 2 молекулы АТФ. Реакция происходит в митохондриях гепатоцитов под действием фермента карбамоилфос-фатсинтетазы I. Карбамоилфосфатсинтетаза II локализована в цитозоле клеток всех тканей и участвует в синтезе гшримидиновых нуклеотидов (см. раздел 10). Карбамоилфосфат затем включается в орнитиновый цикл и используется для синтеза мочевины. В мозге и некоторых других органах может протекать восстановительное аминирование α -кетоглутарата под действием глутаматдегидрогеназы, катализирующей обратимую реакцию. Однако этот путь обезвреживания аммиака в тканях используется слабо, так как глутаматдегидрогеназа катализирует преимущественно реакцию дезаминирования глутамата. Хотя, если учитывать последующее образование глутамина, реакция выгодна для клеток, так как способствует связыванию сразу 2 молекул NH3.
Из мышц и кишечника избыток аммиака выводится преимущественно в виде аланина. Этот механизм необходим, так как активность глутаматдегидрогеназы в мышцах невелика и непрямое дезаминирование аминокислот малоэффективно. Поэтому в мышцах существует ещё один путь выведения азота. Образование аланина в этих органах можно представить следующей схемой (см. схему ниже). Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, основным источником которого служит процесс окисления глюкозы. Мышцы выделяют особенно много аланина в силу их большой массы, активного потребления
Схема глюкозы при физической работе, а также потому, что часть энергии они получают за счёт распада аминокислот. Образовавшийся аланин поступает в печень, где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается, а пируват включается в глюконеогенез. Глюкоза из печени поступает в ткани и там, в процессе гликолиза, опять окисляется до пирувата (рис. 9-13). Образование аланина в мышцах, его перенос в печень и перенос глюкозы, синтезированной в печени, обратно в мышцы составляют глюкозо-аланиновый цикл, работа которого сопряжена с работой глюкозо-лактатного цикла (см. раздел 7). Совокупность основных процессов обмена аммиака в организме представлена на рис. 9-14. Доминирующими ферментами в обмене аммиака служат глутаматдегидрогеиаза и глутаминсинтетаза.
Вопрос 50. Биологическая роль витаминов. Основные причины гиповитаминозов.Биологическая роль- они входят в состав коферментов и простетических групп ферментов, и следовательно используются организмом как строительный материал при синтезе соответствующих небелковых частей ферментов.Гиповитаминоз- специфическое заболевание протекающие в более легкой форме по сравнению в авитаминозами, вызываемым недостаточным содержанием отдельных витаминов в организме.Причины: Экзогенные(связанные с питанием)неправильное приготовление пищи, приготовление пищи с малым кол-вом витаминов, однообразное питание. Эндогенные (связанные с состоянием организма)заболевание ЖКТ и печени, угнетение микрофлоры кишечника, повышенная потребность в витаминах(напр: беременность)
50. Биологическая роль витаминов, основные причины гиповитаминоза. Биологическая роль витаминов. Витамины – это органические соединения с низкомолекулярной структурой. Поступают в организм, в основном, с пищей, так как организм их синтезирует в крайне ограниченных количествах. Виды витаминов: · Водорастворимые витамины (витамины группы В: В1, В2 ,В6, В12, ВС; С; РР; Р; Н). Эти витамины участвуют в образовании различных коферментов. · Жирорастворимые витамины (A1, D2, D3,К и Е) участвуют в определении и поддержании функциональности субклеточных структур и клеточных мембран. При значительном дефиците витаминов все процессы в организме не могут протекать в нормальном режиме, что вызывает нарушения в деятельности органов и их систем. Витамин А (ретинол) нужен для поддержания красивой кожи, волос и всех слизистых, нормальной работы зрительной системы. Без него невозможно гармоничное формирование организма в период отрочества. · Витамин В1 (тиамин) координирует углеводный обмен, поставляющий в организм энергию, поддерживает работу нервной, пищеварительной, дыхательной системы. · Витамин В2 (рибофлавин) отвечает за способность клеток к восстановлению, поэтому при его недостатке даже маленькие кожные трещинки заживают с трудом. Незаменима его функция в процессах окисления и синтеза в организме, а также в поддержании функциональности вегетативного отдела нервной системы. · Витамин В6 (пиридоксин) – участник обмена белков и жиров, стимулирующий использование организмом природных антиоксидантов в виде ненасыщенных жирных кислот. Определенная доля этого витамина образуется микрофлорой кишечника. · Витамин В12 (цианокобаламин) принимает важное участие в процессах кроветворения и белковом обмене. Благодаря этому витамину каротин усваивается организмом, переходя в витамин А. Образуется в толстом кишечнике. · Витамины группы D участвуют в кальциево-фосфорном обмене, поддержке здоровья эндокринных желез. При недостатке - происходит нарушение образования зубов и костей, поражаются мышцы, ухудшается работа пищеварительной, ССС и НС. · Витамин С является важным компонентом окислительно-восстановительных процессов, препятствующим образованию опухолей. Без него не обходятся процессы кроветворения, усвоения железа. Он нужен для поддержки иммунитета. · Витамин Е (токоферола ацетат) – природный антиоксидант, поддерживающий репродуктивные функции. · Витамин РР – один из основных регуляторов обмена веществ, при недостатке которого большинство тканей и органов подвергаются патологическим изменениям. Причины гиповитаминова. · Нехватка витамина в рационе, несбалансированное питание · Разрушение питательных веществ в содержащей их пище вследствие нарушений условий хранения или в результате температурной или иной кулинарной обработки · Действие веществ-антагонистов, которые содержатся в тех или иных продуктах и приводят к разрушению витаминов, нарушению их усвоения (в частности, белок яйца затрудняет усвоение биотина). Гиповитаминоз также может быть обусловлен эндогенными (внутренними причинами): - расстройства работы желудочно-кишечного тракта, приводящие к нарушению процесса всасывания и усвоения витаминов. Часто недостаточное усвоение витаминов бывает вызвано дисбактериозом — нарушением микрофлоры кишечника, которое часто сопровождает длительный прием антибиотиков, прохождение химиотерапии при туберкулезе, онкологических болезнях. Микрофлора участвует в процессе синтеза некоторых витаминов, который происходит внутри организма; ухудшение ее состояния, наличие в ней патогенных микроорганизмов, а также глистные и другие паразитарные инвазии приводят к резкому сокращению запасов витамина в организме. - генетически обусловленные дефекты ферментных систем, транспортных функций, обеспечивающих всасывание и распределение витаминов. - прием некоторых лекарственных препаратов также может вызвать гиповитаминоз. - увеличение потребности человека в витаминах(беременность и кормление, периоды повышенной физической и психической нагрузки, интенсивный рост в подростковом и детском возрасте).
51. В1, В2, В6, РР. Витамин В1.(Тиамин). Используется для синтеза кофермента тиаминдифосфата, необходимого для аэробного распада углеродов. Суточная потребность 2-3мг.Витамин В2.(Рибофлавин). Используется для синтеза коферментов тканевого дыхания-ФАД и ФМН, участвующих в переносе атомов водорода в дыхательной цепи митохондрий. ФАД (флавинадениндинуклеотид) - кофермент, состоящий из двух нуклеотидов соединенных между собой остатками фосфорной кислоты. В состав одного из нуклеотидов входит витамин В2. Совместно в флавиновыми ферментами участвует в переносе атомов водорода в дыхательной цепи митохондрий. ФМН (флавинмононуклеотид) - кофермент, являющийся по строению нуклеотидом, содержащим витамин В2. Совместно с флавиновыми ферментами участвует в переносе атомов водорода в дыхательной цепи митохондрий. Витамин В6. (Пиридоксин). Используется для синтеза кофермента фосфопиридоксаля, участвующего в трансаминировании аминокислот. Суточная потребность 2-3мг. Витамин РР. (Никотинамид). Используется для синтеза коферментов НАД (Никотинамидадениндинуклеотид): необходимого для переноса атомов водорода в дыхательной цепи митохондрий, и НАДФ участвующего в пентозном цикле. Суточная потребность 15-25мг.
Витамины С и Р. Витамин С (Аскорбиновая кислота). Биологическая роль. Участвует в окислительно-восстановительных реакциях. Особенно велика роль витамина С в гидроксировании аминокислот пролина и лизина соответственно в оксипролин и оксилизин при синтезе белка коллагена, а также в синтезе гармона надпочечников. Проявление авитаминоза или гиповитаминоза-цинга. Пищевые источники - Цитрусовые, красный перец, смородина, рябина клюква, квашенная капуста, хвоя. Суточная потребность - 50-100 мг. Витамин Р. Витамин проницаемости (рутин) Биологическая роль. Совместно в витамином С учувствует в окислительно-восстановительных реакциях, снижает проницаемость стенок кровеносных сосудов, обладает антиоксидантными свойствами. Проявление авитаминоза или гиповитаминоза-кровоизлияния Пищевые источники- Цитрусовые, гречиха, красный перец, черноплодная рябина, черная смородина Суточная потребность - Не установлена.
Витамины В12 и В6. Витамин В12 (Цианокобаламин). Биологическая роль-используется для синтеза коферментов,участвующих в переносе метильной группы(-СН3),с последующим включением ее в синтезируемые вещества. Проявление авитаминоза или гиповитаминоза - Анемия Пищевые источники - Печень, почки, мясо, яйца,сыр. Синтезируется микрофлорой кишечника при условии поступления с пищей кобальта. Суточная потребность - 2-3 мкг. Витамин В6. Пиридоксин Биологическая рольиспользуется для синтеза кофермента фосфопиридоксаля, участвующего в трансаминировании аминокислот. Проявление авитаминоза или гиповитаминоза - Дерматид Пищевые источники - печень, почки, мясо, яичный желток. Синтезируется микрофлорой кишечника. Суточная потребность - 2-3 мг. Жирорастворимые витамины. Витамин А(ретинол) Биологическая роль – участвует в восприятии света сетчаткой глаза. Оказывает влияние на барьерную функцию кожи, слизистых оболочек и на проницаемость клеточных мембран. Проявление авитаминоза или гиповитаминоза- Ксерофтальмия (сухость роговой оболочки глаза), кератомаляция (разрушение роговой оболочки),сумеречная или «куриная слепота» Пищевые источники-Жир печени морских рыб, говяжья и свиная печень, яичный желток, морковь. Суточная потребность -2-3 мг. Витамин D (Кальциферол) Биологическая роль - участвует во всасывании в кишечнике ионов Са, их транспорте кровью и во включении их в состав костной ткани и в процессе окостенения Проявление авитаминоза или гиповитаминоза - рахит. Пищевые источники: Жир печени морских рыб, сливочное масло, растительные масла,яйца, молоко. Суточная потребность - 13-25 мкг для детей и для беременных, 7-12 мкг для взрослых. Витамин Е.(токоферол). Биологическая роль-является главным антиоксидантом организма, предохраняющим от окисления полиненасыщенные жирные кислоты, входящие в биологические мембраны. Проявление авитаминоза или гиповитаминоза: У экспериментальных животных- бесплодие, мышечная дистрофия. Пищевые источники- злаки, растительные масла, мясо сливочное масло. Суточная потребность-5-10мг. Витамин К (Филлюхинон). Биологическая роль - участвует в синтезе некоторых факторов свертывания крови(в том числе протромбина) Проявление авитаминоза или гиповитаминоза - повышенная кровоточивость Пищевые источники- Печень, шпинат, морковь, капуста. Синтезируется микрофлорой кишечника Суточная потребность -100 мкг.
55. Общие механизмы действия гормонов. 3. Влияют на проницаемость клеточных мембран по отношению к определенным химическим соединениям. В результате этого в клетки поступает больше или меньше субстратов для ферментативных реакций, что сказывается на скорости химических процессов. По химическому строению делятся на: 1. Гормоны белковой природы (белки и полипептиды): гормоны гипоталамуса, гормоны гипофиза, кальцитонин щитовидной железы, гормон паращитовидных желез, гормоны поджелудочной железы; 2. Гормоны - производные аминокислоты тирозина: йодсодержащие гормоны щитовидной железы, гормоны мозгового слоя надпочечников; 3. Гормоны стероидного строения: гормоны коры надпочечников, гормоны половых желез.
56. Гормоны гипоталамуса и гипофиза. Гипоталамус. Либерины (рилизинг факторы)– Химическая природа гормона - белок Стимулирует выделение в кровь гормонов передней доли гипофиза. Статины (ингибирующие факторы)– Химическая природа гормона- белок Тормозят выделения в кровь гормонов передней доли гипофиза.   Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|