|
|
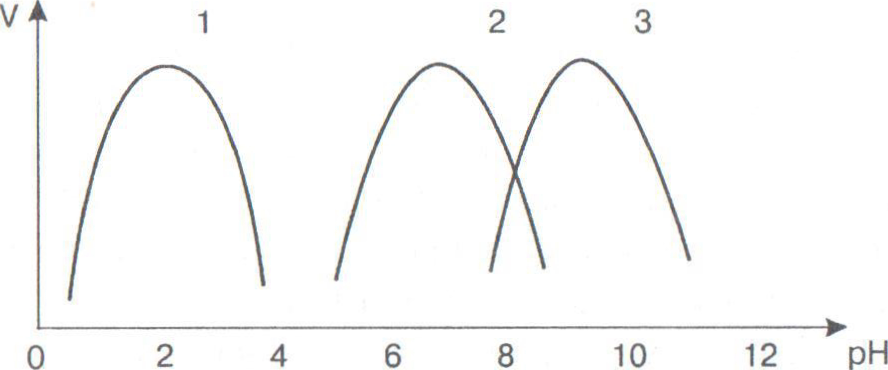
Зависимость скорости ферментативной реакции от pH
11) Ингибиторы и активаторы ферментов Ингибиторы (I) - это химические соединения (обычно низкомолекулярные), которые, находясь в низких концентрациях, избирательно тормозят определенные ферментативные реакции. При этом ингибитор всегда присоединяется к ферменту с образованием фермент-ингибиторного комплекса. Фермент, связанный с ингибитором, теряет свою каталитическую активность.Если связи между ферментом и ингибитором прочные, то действие ингибитора носит необратимый характер, и торможение нарастает во времени вплоть до полного прекращения ферментативной реакции:
Такие ингибиторы называются необратимыми. Примером необратимых ингибиторов являются фосфорорганические соединения (табун, зарин, дихлофос, карбофос и др.), избирательно и необратимо угнетающие важнейший фермент организма холинэстеразу, участвующую в передаче нервных импульсов.Если ингибитор присоединяется к ферменту за счет непрочных связей, то торможение фермента является обратимым и не зависит от времени:
Ингибиторы такого типа называются обратимыми. В свою очередь, обратимые ингибиторы делятся на две разновидности: конкурентные и неконкурентные. Конкурентные ингибиторы присоединяются к активному центру фермента, т.е. к тому же участку поверхности фермента, что и субстрат. Поэтому между ингибитором и субстратом идет конкуренция за присоединение к активному центру. Занимая активный центр, ингибитор тем самым препятствует образованию фермент-субстратного комплекса - первой стадии ферментативного катализа. Конкурентные ингибиторы обычно по строению похожи на субстрат. Неконкурентные ингибиторы присоединяются к ферменту вне активного центра. Этот участок поверхности фермента называется аллостерический центр (т.е. находящийся в другом месте по сравнению с активным центром).Присоединение неконкурентного ингибитора к аллостерическому центру вызывает неблагоприятное изменение пространственной структуры (конформации) всей молекулы фермента, в т.ч. и активного центра. В результате каталитические свойства фермента снижаются. Неконкурентные ингибиторы участвуют в регуляции скорости ферментативных реакций, протекающих в организме. В роли неконкурентных ингибиторов могут быть гормоны, конечные и промежуточные продукты ферментативных реакций, ионы металлов, лекарственные вещества. Активаторы - вещества, избирательно повышающие скорость определенных ферментативных реакций. Активаторы, подобно неконкурентным ингибиторам, присоединяются обратимо к аллостерическому центру фермента. Но в этом случае изменение конформации фермента будет благоприятным для функционирования активного центра, что приводит в итоге к повышению скорости ферментативной реакции. В организме активаторами являются некоторые гормоны, промежуточные продукты метаболических процессов, ионы металлов, а также лекарственные препараты.
12) Классификация и индексация ферментов Современная классификация ферментов основывается на характере химической реакции, катализируемой ферментом. Все ферменты делятся на шесть классов в зависимости от типа катализируемой реакции: I класс - оксидоредуктазы - ферменты, катализирующие окислительно-восстановительные реакции. Схематично действие ферментов I класса можно записать: А+в —>C + D II класс - трансферазы - ферменты, катализирующие перенос химических группировок с молекулы одного вещества на молекулу другого: АВ + С —>А + ВС III класс - гидролазы - ферменты, расщепляющие химические связи путем присоединения воды, т.е. путем гидролиза: АВ + н20 > А-Н + В-ОН IV класс - лиазы - ферменты, катализирующие расщепление химических связей без присоединения воды: АВ—>А+В V класс - изомеразы - ферменты, катализирующие изомерные превращения, т.е. перенос отдельных химических групп в пределах одной молекулы: А—>в VI класс - синтетазы - ферменты, катализирующие реакции синтеза, протекающие за счет энергии гидролиза АТФ: А+В ^ АВ АТФ + Н20 ^ ^ АДФ + Н3Р04(изображение чтобы посмотреть последнюю формулу)
В основе подразделения ферментов на подклассы и на подподкласе лежит тип химической связи или химическое строение группировки ■ убстрата, на которую действует фермент. С классификацией ферментов тесно связана их индексация (нумерация). Индекс (шифр) каждого фермента состоит из четырех чисел, разделенных точками, и составляется по следующему принципу: перцам цифра индекса указывает, к какому из шести классов принадлежит манный фермент. Второе и третье числа индекса обозначают соответственно порядковые номера подклассов и подподклассов. Четвертое число индекса - порядковый номер индивидуального фермента внутри | своего подподкласса. Например, фермент слюны амилаза, расщепляющий крахмал, имеет индекс 3.2.1.1, что свидетельствует о принадлежности этого фермента N классу гидролаз
Вопрос 13. Регуляция скорости ферментативных реакций. К основным механизмам регуляции скорости ферментативных реакций можно отнести следующие: 1- Изменение скорости синтеза ферментов. Ферменты, являясь белками, постепенно разрушаются, и поэтому в клетках постоянно синтезируются новые ферментные белки. При повышении скорости образования новых ферментов их содержание в клетках возрастает. При этом скорость ферментативных реакций увеличивается, так как зависимость скорости реакции от концентрации фермента носит прямолинейный характер. При уменьшении выработки ферментов их концентрация в клетках постепенно снижается, что сопровождается падением скорости катализируемых ими реакций.Изменение скорости синтеза ферментов обычно происходит под влиянием гормонов, которые ускоряют или замедляют первый этап синтеза ферментных белков - считывание генетической информации – транскрипцию. Поэтому данный механизм регуляции часто называется регуляцией на генетическом уровне. 2- Модификация ферментов. Под модификацией понимается незначительное изменение химического состава молекулы фермента, приводящее к изменению каталитической активности.Модификация может быть обратимой и необратимой.В первом случае изменение строения фермента носит обратимый характер и фермент может многократно менять свою активность. Например, некоторые ферменты содержат в своей молекуле остаток фосфорной кислоты. При отщеплении фосфатного остатка у одних ферментов каталитическая активность снижается; при включении его снова в состав ферментов активность их возрастает. У других же ферментов фосфорилированная форма (содержащая фосфат), наоборот, малоактивна. Отщепление фосфорной кислоты от таких ферментов ведет к повышению их активности.Примером необратимой модификации служит активация пищеварительных ферментов, действующих на пищевые белки. Так, в составе желудочного сока содержится белок пепсиноген, не обладающий каталитической активностью. В полости желудка под действием соляной кислоты от этого белка отщепляется небольшой полипептид и образуется очень активный фермент, расщепляющий в пищевых белках пептидные связи – пепсин. (Белки, являющиеся предшественниками ферментов, называются проферментами). 3- Изменение конформации ферментов. Изменение конформации, т.е. пространственного строения ферментного белка происходит под действием неконкурентных ингибиторов и активаторов, которыми могут быть гормоны, промежуточные и конечные продукты ферментативных реакций, ионы металлов, лекарства (см. выше). Изменение конформации ферментов возможно и при изменении условий среды, например, при повышении или снижении кислотности.
Вопрос 14. Общая характеристика обмена веществ. Пищеварение и Метаболизм. Общая характеристика обмена веществ Обмен веществ и энергии – это обязательное условие существования живых организмов. 1-В процессе катаболизма преобладают реакции окисления. 2- Катаболизм протекает с потреблением кислорода. 3- В процессе катаболизма освобождается энергия, примерно половина которой аккумулируется в форме химической энергии аденозинтрифосфата (АТФ). Другая часть энергии выделяется в виде тепла. Анаболизм включает разнообразные реакции синтеза.Анаболизм характеризуется следующими особенностями: ·Для анаболизма типичны реакции восстановления. · В процессе анаболизма происходит потребление водорода. Обычно используются атомы водорода, отщепляемые от глюкозы и переносимые коферментом НАДФ · Анаболизм протекает с потреблением энергии, источником которой является АТФ. Основное назначение метаболизма: · Одновременное протекание реакций катаболизма и анаболизма приводит к обновлению химического состава организма, что является обязательным условием его жизнедеятельности. · В случае преобладания анаболизма над катаболизмом происходит накопление химических веществ в организме и, в первую очередь, белков. Накопление белков в организме - обязательное условие его роста и развития. · Обеспечение энергией (в форме молекул АТФ) всех потребностей организма.
Вопрос 15. Строение и биологическая роль АТФ. Аденозинтрифосфат (АТФ) является нуклеотидом. В состав молекулы АТФ входят азотистое основание - аденин, углевод - рибоза и три остатка фосфорной кислоты (аденин, связанный с рибозой, называется аденозином). Особенностью молекулы АТФ является то, что второй и третий остатки фосфорной кислоты присоединяются связью, богатой энергией. Такая связь называется высокоэнергетической или макроэргической и обозначается знаком ~. Соединения, имеющие макроэргические связи, обозначаются термином «макроэрги» Структурная формула АТФ имеет следующий вид: В упрощенном виде строение АТФ можно отразить схемой: аденозин При использовании АТФ в качестве источника энергии обычно происходит отщепление путем гидролиза последнего остатка фосфорной кислоты: АТФ + Н2О -> АДФ + Н3РО4 + Q (энергия) Главными потребителями энергии АТФ в организме являются: · Реакции синтеза · Мышечная деятельность · Транспорт молекул и ионов через мембраны (например, всасывание веществ из кишечника, образование мочи в почках, формирование и передача нервного импульса и др.).Биологическая роль АТФ заключается в том, что это вещество является универсальным аккумулятором энергии, своего рода энергетической «валютой» клетки.Основным поставщиком АТФ является тканевое дыхание - завершающий этап катаболизма, протекающий в митохондриях всех клеток, кроме красных клеток крови (эритроцитов).
16. Ферменты тканевого дыхания. Тканевое дыхание - основной способ получения АТФ, используемый всеми клетками организма (кроме эритроцитов). Ферменты тканевого дыхания: 1) Никатинонамидные дегидрогеназы: отнимают 2 атома водорода от окисляемого субстрата временно присоединяют их к своему никатинамидадениндинуклеотиду (НАД). При этом присоединении НАД превращается в свою восстановленную форму: АН2+НАД=А + НАД*Н2, где А - окисляемый субстрат. 2) Флавиновые дегидрогеназы: отщепляют 2 атома водорода от НАД*Н2 и временно присоединяет их к флавинмононуклеотиду (ФМН). Является мононуклеотидом, содержащим рибофлавин (витамин В2). В результате образуется восстановленная форма кофермента - ФМН*Н2: НАД*Н2 + ФМН = НАД + ФМН*Н2. В некоторых случаях флавиновые дегидрогеназы отнимают атомы водорода непосредственно от окисляемого вещества. Они используют кофермент ФАД (флавинадениндинуклеотид). Отнимаемые от окисляемого вещества атомы водорода также присоединяются к флавину: АН2 + ФАД = А + ФАД*Н2. 3) Цитохромы: участвуют только в переносе электронов. По строению похожи на одну из субъединиц гемоглобина, состоит из полипептида и гема. В состав гема входит железо с переменной валентностью, что и отличает от гема эритроцитов. Именно эта особенность железа даёт возможность переносить электроны: Fe3+ + e = Fe2+ С помощью цитохромов b, c, a, a3 электрроны от восстановленных коферментов ФМН*Н2 или ФАД*Н2 передаются на молекулярный кислород О2, который при этом переходит в активную, анионную форму О2-. Далее этот кислород связывается с ионами водорода, который отщепляется от ФМН*Н2 или ФАД*Н2. Это приводит к образованию воды. Некоторые субстраты имеют более высокий редокс-потенциал, чем НАД. Поэтому они не могут окисляться никотинамидными дегидрогеназами. В этом случае отнятие атомов водорода происходит флавиновыми дегидрогеназами. В таком случае образуется только 2 молекулы АТФ.
17. Схема дыхательной цепи. Синтез АТФ в процессе тканевого дыхания.
Движение электронов сопровождается выделением энергии. Около половины энергии аккумулируется в макроэнергетических связях АТФ. Другая часть энергии рассеивается в виде тепла. Количество выделяемой энергии зависит от величина редокс-потенциала, поэтому синтез АТФ происходит на участках цепи с большой разностью редокс-потенциала: v При переносе электронов с НАД*Н2 на ФМН. v С цитохрома b на цитохром с. v С цитохрома а на а3. Итого при переносе двух атомов водорода на кислород синтезируется 3 молекулы АТФ. Некоторые субстраты имеют более высокий редокс-потенциал, чем НАД. Поэтому они не могут окисляться никотинамидными дегидрогеназами. В этом случае отнятие атомов водорода происходит флавиновыми дегидрогеназами. В таком случае образуется только 2 молекулы АТФ. В сутки в организме образуется не менее 40 кг АТФ, у спортсменов ещё больше. Чем больше клетка использует АТФ, тем больше скорость окислительного фосфорилирования. Причина: активатором ферментов процесса тканевого дыхания является избыточный уровень АДФ (аденозиндифосфат), который выделяется при использовании АТФ. Дыхательный ансамбль - отдельные скопления ферментов тканевого дыхания на внутренней мембране митохондрии.
18. Анаэробное окисление. В некоторых случаях отнятие атомов водорода происходит в цитоплазме и присоединение происходит не к атомам кислорода, а к другим веществам. Наиболее часто акцептором является ПВК (пировиноградная кислота), возникающая при распаде аминокислот и углеводов. В результате ПВК превращается в лактат (молочную кислоту). Этот процесс происходит без потребления кислорода, т.е. анаэробно. За счёт выделяющейся энергии в цитоплазем происходит синтез АТФ, который называется анаэробное фосфорилирование.
19 вопрос Микросомальное окисление Происходит когда при окислении атомы кислорода включаются в молекулы окисляемых веществ. Такое окисление протекает в мембранах цитоплазматической сети и носит название микросомальное окисление. А когда в молекуле окисляемого субстрата включается кислород возникает гидроксилиная группа (-ОН),а само окисление называют гидросилированием. В нем принимает участие витамин С. Биологическая роль микросомального окисления: 1)включение атомов кислорода в синтезируемого вещества 2)обезвреживание различных токсических соединений, поступающих в организм из вне или образующихся в процессе метаболизма. Включение кислорода в молекулу яда и снижает его токсичность, делая его более водорастворимыми и облегчая выведения из организма почками.
20 вопрос. Cвободнорадикальное окисление Незначительная часть кислорода,поступающего из воздуха в организм превращается в активные формы(О2-,НО2-,НО,)называют свободными радикалами или оксидантами
Перекись водорода Участок молекулы жирной кислоты Подвергшейся СРО
I I H – H – C – C– I I I I O – O O – O
Далее в этом месте происходит расщепление жирной кислоты. Перекисное возникновение липидов(ПОЛ) –это свободнорадикальное окисление жирных кислот,входящих в состав липидов, возникших в связи перекисной группировки. Образование свободных радикалах кислорода в тканях организма происходит постоянно. За счет этого процесса осуществляется обновление липидного слоя биологических мембран. Свободные радикалы выполняют также защитную функцию,окисляя различные чужеродные вещества, поступающие в организм из вне, в том числе мембранные белки и липойды патогенных микроорганизмов. В физиологических условиях свободнорадикальное окисление протекает с низкой скоростью, так как ему противостоит защитная антиоксидантная система организма, предупреждающая накопление свободных радикалов кислорода и ограничивающая тем самым скорость СРО.Главным компонентом антиоксидантной системы является витамин Е (токоферол). Активные формы кислорода могут возникать в процессе тканевого дыхания. Для нормального течения тканевого дыхания необходимо одновременное присоединение к молекуле кислорода четырех электронов. В этом случае образуются две молекулы воды. Иногда к молекуле кислорода присоединяется два или один электрон и возникает перекись водорода и раксид-анион кислорода, которые очень токсичны для клеток, так как являются сильными окислителями и повреждают биомембраны Для защиты этих опасных соединений в состав антиоксидантной системы входят специальные ферменты. Под действием фермента супероксиддисмутазы супероксид-анион превращается в перекись водорода
2О2+ 2Н→Н2О2+ О2 Далее перекись водорода разрушается ферментом католазой.
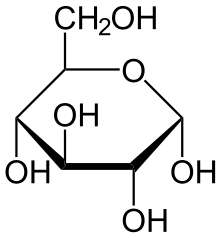
21вопрос. Строение и биологическая роль глюкозы. Важнейшим природным углеродом является глюкоза,которая может находиться как и в свободном виде (моносахарид) так и в составе олигосахаров (сахароза,лактоза и др.) и полисахоридов(клечастка,крахмал,гликоген) Формула глюкозы C6H12O6
Свободная глюкоза в организме человека в основном находится в крови, где ее содержание довольно постоянно и колеблется в узком диапазоне от 3,9 до 6,1 ммоль/л(70-110мг%,или 07-1,1г/л).Глюкоза в организме в первую очередь выполняет энергетическую функцию и является главным источником энергии.
  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
 E + I EI
E + I EI
 E + I EI
E + I EI
 На всём протяжении схемы видно передвижение электронов. оно вызвано тем, что в дыхательной цепи все участники расположены по увеличению своего окислительно-восстановительного потенциала, или редокс-потенциала. Это способность вещества принимать и удерживать электроны. Электроны переносятся от вещества с низким потенциалом к веществу с более высоким.
На всём протяжении схемы видно передвижение электронов. оно вызвано тем, что в дыхательной цепи все участники расположены по увеличению своего окислительно-восстановительного потенциала, или редокс-потенциала. Это способность вещества принимать и удерживать электроны. Электроны переносятся от вещества с низким потенциалом к веществу с более высоким.