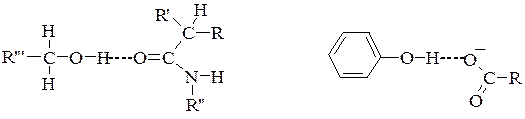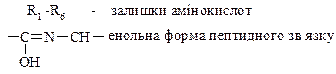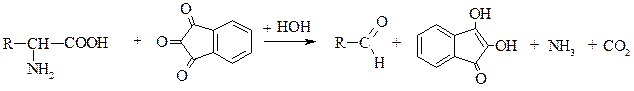|
|
Значення білків у життєдіяльності людиниТа чи інша молекула білка має генетично закодований чітко визначений амінокислотний склад, специфічні фізико-хімічні властивості та відповідну специфічну біологічну функцію у живій клітині, що обумовлює цільове призначення білка. Ні один клас біологічно-активних речовин не виконує стільки функцій як білки. Структурна функція Білки, що виконують опорну функцію, займають по кількості перше місце серед інших білків тіла людини. Серед них найважливішу роль відіграють фібрилярні білки, зокрема колаген сполучної тканини, кератин волосся, нігтів, шкірі, еластин стінок судин, і т.п. Білки входять до складу всіх клітинних органел і надають їм форму: мембранних (плазмалема, ядерна оболонка, ендоплазматичний ретикулум (ЕР), комплекс Гольджі, лізосоми, пероксисоми, вакуоль, мітохондрії, пластиди) і немембранних – хромосоми, рибосоми, клітинний центр (центріолі), війки та джгутики, мікрофіламенти. Каталітична функція Ферменти – це біологічні каталізатори білкової природи. До 1995 р. було ідентифіковано більш ніж 3400 ферментів. За участю ферментів у клітинах одночасно проходить багато різних хімічних реакцій, які забезпечують синтез і розпад різноманітних сполук з досить високою швидкістю при температурі 37°С та атмосферному тиску. В 1982 році з'ясувалося, що деякі РНК теж мають каталітичну активність. Їх називають рибозимами. Захисна функція Основну функцію захисту в організмі виконує імунна система, котра забезпечує синтез специфічних захисних білків-антитіл у відповідь на надходження в організм бактерій, токсинів, вірусів або чужорідних білків. Антитіла – це білки. Імуноглобуліни «склеюють» антигени й утворюється преципітат. Захисна функція білків проявляється також і у здатності білків плазми крові (фібриногену) до згортування. 4) Регуляторна функція На рівні організму деякі гормони мають білкову природу і регулюють обмін речовин. Наприклад, інсулін – гормон підшлункової залози регулює перехід глюкози через плазмалему. При недостатній секреції інсуліну розвивається важке захворювання – цукровий діабет. Соматотропін – гормон росту. Утворюється в передній частині гіпофіза і відіграє важливу роль в забезпеченні анаболічних процесів, сприяє синтезу білків та нуклеїнових кислот. Там же утворюється й адренокортикотропний гормон (АКТГ). Він діє на кору наднирок, регулюючи синтез стероїдних гормонів. Тиротропін забезпечує нормальне функціонування щитоподібної залози. 5) Транспортна функція Для нормальної життєдіяльності організму необхідне постійне забезпечення його органів і тканин поживними речовинами. Ці сполуки переносяться з током крові сполуками білкової природи. Так дихальна функція крові, зокрема перенос кисню до тканин і на зворотному шляху транспорт вуглекислого газу до легень, здійснюється за рахунок білку еритроцитів гемоглобіну. Білок трансферин здійснює транспорт заліза. У транспорті ліпідів приймають участь альбуміни сироватки крові. Системи мембранних білків –пермеази переносять полярні сполуки через мембрану як за, так і проти градієнта концентрації. 6) Енергетична функція В організмі людини 11 з 20 амінокислот, що входять до складу білків, розщеплюються з виділенням енергії. Це – замінні амінокислоти. Вони можуть бути синтезовані в клітині з продуктів розщеплення вуглеводів і ліпідів. При розщепленні 1г білка виділяється 17,7кДж енергії. За рахунок білків організм людини одержує 10 – 15% енергії. Резервна (запасаюча) функція. Цю функцію виконують білки, які запасаються в якості джерела енергії та речовин в насінні рослин та яйцеклітинах тварин для розвитку зародка і вигодовування дитинчат. Наприклад, альбумін – яєчний білок, гліадин – білок зерен пшениці, казеїн – білок молока, який також виконує живильну функцію. Ряд інших білків використовується в якості джерела амінокислот, з яких потім синтезуються інші біологічно-активні сполуки, що регулюють метаболічні процеси організму. Буферна функція. Будь-який білок – амфотерний поліелектроліт. Білки сприяють підтримці визначених значень рН у різних відсіках клітини, забезпечуючи цим компартменталізацію (цілісність) організму. 9)Трансформація енергії. Білки сітчатки ока родопсин і ретинон трансформують світлову енергію в електричну. Актино-міозинові комплекси в м'язах перетворюють енергію хімічних зв'язків у механічну. Механічна функція. Білки беруть участь у забезпеченні різних форм механічного руху – переміщенні клітин в організмі, скороченні і розслабленні м‘язів, роботі внутрішніх органів – серця, легень, шлунка. Ці процеси здійснюються за участю актину, міозину тропоміозину та інших білків. Будова білків. Амінокислоти Білки побудовані з L-амінокислот. Амінокислоти – органічні карбонові кислоти,в молекулах яких один або декілька атомів водню заміщені аміногрупою. · Усі амінокислоти, що входять до складу білків, є α-амінокислотами, оскільки їх аміногрупа перебуває у α-положенні, тобто поряд з карбоксильною групою. · Усі амінокислоти, за винятком гліцину, мають асиметричний атом вуглецю (тобто атом вуглецю з чотирма різними замісниками) і тому є оптично активними сполуками. Амінокислоти, що містяться в білках, належать до L-ряду. L-конфігурація – аміногрупа розташована зліва, D-конфігурація – аміногрупа розташована справа. · Амінокислоти – амфотерні електроліти, оскільки аміногрупа має основні властивості, а карбоксильна – кислотні. У водних розчинах в інтервалі рН 4 – 9 амінокислоти існують переважно у вигляді біполярних йонів, які називаються цвітер-іонами.
Скорочено амінокислоти позначають трьома літерами або однією літерою. Наприклад, аланін – Ala (A), аргінін – Arg (R). Існує декілька класифікацій амінокислот: · За хімічною структурою амінокислоти поділяють на: аліфатичні, гетероциклічні, ароматичні. · Біологічна. Вищі рослини та більшість мікроорганізмів синтезують 20 амінокислот. Амінокислоти можна поділити на три групи. Перші – синтезуються в організмі, вони називаються замінними; другі – в недостатній кількості синтезуються в організмі – напівзамінні; треті – надходять з їжею - незамінні (таких кислот 8: Val, Leu, Ile, Phe, Trp, Thr, Lys, Met). У живих організмах нараховується 21 протеїногенна амінокислота. Замінні амінокислоти здатні замінювати одна іншу в раціоні, оскільки вони перетворюються одна в одну або синтезуються з проміжних продуктів вуглеводного або ліпідного обміну. Відсутність в їжі хоча б однієї незамінної амінокислоти викликає негативний азотистий баланс, порушення діяльності центральної нервової системи, зупинку росту і важкі клінічні наслідки типу авітамінозу. Недостатня кількість однієї незамінної амінокислоти призводить до неповного засвоєння інших. Дана закономірність підкоряється закону Лібіха, за яким розвиток живих організмів визначається тією незамінною речовиною, яка присутня в найменшій кількості. Залежність функціонування організму від кількості незамінних амінокислот використовується при визначенні біологічної цінності білків хімічними методами. Найбільш широко використовується метод X. Мітчела і Р. Блоку (Mitchell, Block, 1946), відповідно до якого розраховується показник амінокислотного скору (А.С.). Скор виражають у відсотках або безрозмірною величиною, що представляє собою відношення вмісту незамінної амінокислоти (А.К.) в досліджуваному білку до її кількості в еталонному білку. При розрахунку скора у відсотках формула виглядає наступним чином:
Амінокислотний склад еталонного білка збалансований та ідеально відповідає потребам організму людини в кожній незамінній кислоті, тому його ще називають «ідеальним». Амінокислота, скор якої має найнижче значення, називається першою лімітуючою амінокислотою. Значення скора цієї амінокислоти визначає біологічну цінність і ступінь засвоєння білків. Наприклад, А.С. курячого яйця -1; коров‘ячого молока - 0,95; рису - 0,67; сої - 0,55; пшениці - 0,53. Зазвичай розраховують скор для трьох найбільш дефіцитних амінокислот: лізин, метіонін, триптофан 3:3:1. Співвідношення суми незамінних амінокислот до суми замінних не повинно бути нижчим 0,4. · Класифікація амінокислот, що входять до складу білків, за принципом полярності (неполярності) радикала (бічного ланцюга). Амінокислоти з неполярними або гідрофобними радикалами: Аліфатичні амінокислоти – аланін, валін, лейцин, ізолейцин, метіонін. Ароматична – фенілаланін. Гетероциклічна – триптофан. Імінокислота – пролін.
Аланін Валін Лейцин
Ізолейцин Метіонін
Фенілаланін Триптофан Пролін Амінокислоти з полярними, незарядженими радикалами: Гліцин. Гідроксиамінокислоти – серин, треонін, тирозин. Амінокислота з сульфгідрильною групою – цистеїн. З амідною групою – аспарагін, глютамін.
Гліцин Серин Треонін
Цистеїн Аспарагін Глютамін
Тирозин 3. Амінокислоти з негативно зарядженими радикалами:
Аспарагінова кислота Глютамінова кислота 4. Амінокислоти з позитивно зарядженими радикалами:
З амінокислот будуються пептиди та білки. Цей процес нагадує нанизування намистинок на нитку. При цьому можливі найрізноманітніші сполучення амінокислот. Послідовність амінокислот у «намисті» називається первинною структурою білка. Оскільки намистинкою може 21 амінокислота, то навіть для коротких білків існує величезна кількість можливих варіантів первинної структури. Наприклад, існує більш 1012 варіантів зібрати білок довжиною усього з 10 амінокислот! 3.2.1. Способи добування амінокислот Основні методи добування амінокислот можна поділити на групи: 1) Гідроліз білків – кислотний, лужний, ферментативний. 2) Хімічний синтез. В результаті отримують оптично неактивну рацемічну суміш L- i D-форми. Потім розділяють рацемат за допомогою іммобілізованих ферментів. 3) Мікробіологічний синтез: триптофан, лізин Майже всі α-амінокислоти, що надходять з травного тракту людини в кров'яне русло організму, зазнають ряд загальних перетворень, призначення яких полягає в забезпеченні пластичним матеріалом процесу синтезу білків і пептидів та здійсненні дихання з утворенням АТФ (рис. 6). В основі таких перетворень лежать реакції дезамінування, трансамінування та декарбоксилю-вання амінокислот.
Рис. 6. Перетворення амінокислот в організмі. 3.2.2. Способи зв¢язку амінокислот у молекулі білку. 1) Ковалентні зв’язки у молекулі білка можуть бути двох типів – пептидні та дисульфідні. У білках наявний кетоімідний тип зв'язку, який був названий пептидним. Пептидний зв'язок утворюється внаслідок взаємодії карбоксильної групи однієї амінокислоти з аміногрупою другої амінокислоти. Сполуки, які при цьому утворюються, називаються пептидами. Схематично цей процес можна показати так:
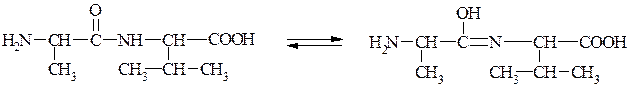
Дослідженнями встановлено, що пептидний зв'язок є проміжним між подвійним і простим (одинарним) зв'язком. Так, відстань між атомами Карбону і Нітрогену (С—N) в пептидному зв'язку дорівнює 0,132 нм, тоді як довжина одинарного зв'язку між Карбоном і Нітрогеном становить 0,147 нм, а подвійного — 0,125 нм. Ця особливість пептидного зв'язку зумовлює таутомерне (кето-енольне) перетворення:
Пептидний з'вязок є основою утворення поліпептидних ланцюгів, з яких побудовано молекули білка. Ковалентні зв'язки між залишками двох цистеїнів (дисульфідні містки). Цей досить міцний ковалентний зв'язок утворюється внаслідок відщеплення атомів Гідрогену від сульфгідрильних груп двох амінокислотних залишків цистеїну. Дисульфідний зв'язок, як і водневий, може бути внутрішньоланцюговим і міжланцюговим. Велика кількість дисульфідних зв'язків є також в білках сполучної і покривної тканин та в білках, які мають високу біологічну активність. Дисульфідні зв'язки мають важливе значення в формуванні третинної структури білків. Руйнування цих зв'язків призводить до дестабілізації даного рівня структури і втрати білком його біологічної активності. 2) Іонні (електростатичні) взаємодії між протилежно зарядженими амінокислотними залишками. Між залишками моноамінодикарбонових та діаміномонокарбонових кислот. Наприклад, позитивно заряджена e-аміногрупа лізину (–NН3+) притягується негативно зарядженою карбоксильною групою – (–СОО–) глютамінової або аспарагінової кислоти. Виникає електростатична взаємодія. Ці взаємодії приймають участь в утворенні третинної та четвертинної структури. 3) Водневі зв'язки – це слабка електростатична взаємодія між одним електронегативним атомом (наприклад, Оксигеном чи Нітрогеном) та атомом Гідрогену, що ковалентно-зв'язаний з другим електронегативним атомом. Беруть участь всі амінокислоти, що у радикалі мають гідроксильні, амідні або карбоксильні групи.У молекулах білка водневий зв'язок найчастіше утворюється при взаємодії атома Гідрогену імінної групи залишку однієї амінокислоти з атомами Оксигену карбонільної групи залишку іншої амінокислоти:
Водневий зв'язок у молекулі білка може бути внутрішньоланцюговим (з’єднує окремі витки однієї спіралі) і міжланцюговим (з’єднує різні поліпептидні ланцюги). У нативних білках цей тип зв'язку може утворюватися не лише між Гідрогеном і Оксигеном пептидних груп, а й між іншими функціональними групами поліпептидного ланцюга. Водневий зв’язок досить слабкий але грає важливу роль в утворенні вторинної, третинної та четвертинної структур.
Водневий зв'язок між атомом Гідрогену гідроксильної групи і атомом Оксисену карбонільної групи пептидного зв’язку.
Водневий зв'язок між атомом Оксисену і атомом Гідрогену двох гідроксильних груп або гідроксильної та аміногруп. 4) Гідрофобні взаємодії. Утворюються внаслідок міжмолекулярної взаємодії (сил Ван-дер Ваальса) між неполярними алкільними радикалами у водному середовищі. Залишок лейцину
Залишок валіну Групи, що можуть утворювати водневі зв'язки з молекулами води, називаються гідрофільними. До них відносяться гідрокси- і аміногрупи, карбонільна, карбоксильна, амідна групи і т.п. Молекули або частини молекули, які не здатні утворювати водневі зв'язки з водою називаються гідрофобними. До гідрофобних груп відносяться алкільні й ароматичні радикали. Як правило, гідрофобні сполуки – або погано, або зовсім нерозчинні, гідрофільні добре розчинні у воді. Особливо важливим є дослідження послідовності розміщення залишків амінокислот у поліпептидних ланцюгах, тобто первинної структури білка. Остання визначає трьохмірну структуру поліпептидних ланцюгів, яка називається конформацією білка. 3.2.3. Рівні просторової організації біополімерів При розгляді просторової організації біополімерів виділяють чотири рівні: первинну, вторинну, третинну і четвертинну структуру. Хоча такий поділ значною мірою умовний, він досить корисний для систематизації знань про будову біополімерів. Первинна структура білка -це послідовність розташування амінокислотних залишків у поліпептидному ланцюзі. Первинною структурою називають порядок, у якому мономерні одиниці зв'язані між собою ковалентними зв'язками. На Землі не було, немає і не буде двох людей з цілком однаковим набором білків. Вторинна структура білка – це просторово упорядкована будова поліпептидних ланцюгів, обумовлена водневими зв'язками між групами С=О і N–Н різних амінокислот. Цей процес відбувається не хаотично, а у відповідності з програмою, що закладена у первинній структурі. Детально досліджені дві основні конфігурації - a-спіраль (правозакручена) та b-конформація. Одним з основних елементів вторинної структури білків є a-спіраль. Це – права спіраль (запропонована Лайнусом Полінгом і Робертом Кору), що містить 3,6 амінокислотні залишки на витокі з періодом повторюваності (кроком) 5,4Ǻ або 0,54 нм. Поліпептидний кістяк утворює щільні витки навколо довгої осі молекули, тоді як бічні радикали виступають назовні. Спіраль утримується внутрішньомолекулярними водневими зв'язками між групами N–Н одного пептидного зв'язку і атомом Оксигену групи С=О, що належить іншому пептидному зв'язку, розташованому через чотири амінокислотні залишки над першим в наступному витку спіралі. Період ідентичності, тобто довжина відрізка спіралі, яка повністю повторюється по ходу її, становить 2,7 нм і включає 18 залишків амінокислот. Деякі амінокислоти (аланін, валін, лейцин, метіонін, фенілаланін, тирозин, триптофан, гістидин тощо) здатні до утворення a-спіралі, особливо коли вони розміщені поряд у поліпептидному ланцюгу. Інші амінокислоти такі, як лізин, аргінін, гліцин, серин, треонін, аспарагінова і глутамінова кислоти, сприяють дестабілізації a-спіралі. Разом з цим окремі амінокислоти, зокрема пролін і оксипролін, просторово в спіральну структуру не вкладаються. На цих ділянках напрям поліпептидного ланцюга змінюється на 103°, і спіральна структура порушується. Будову a-спіралі схематично показано на рис. 7.
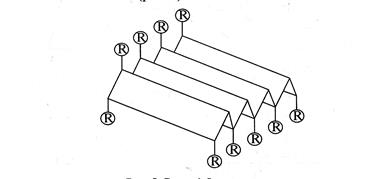
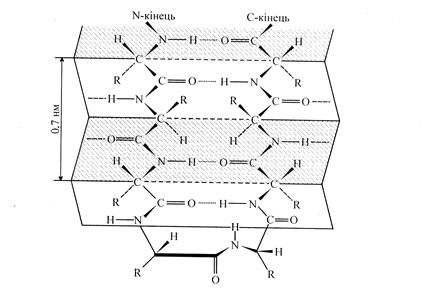
Рис. 7. Вторинна структура білка. α-спіраль Другим елементом вторинної структури білків є b-конформація. Остов поліпептидного ланцюга в b-конформації виглядає таким чином, що має вже не спіральну, а зиґзаґоподібну, гофровану форму. Вважають, що b-структура може існувати у двох різновидностях. Перша дістала назву антипаралельного складчастого шару. Вона утворена антипаралельними поліпептидними ланцюгами, тобто ланцюгами, N -кінці яких напрямлені в протилежні боки.
Рис. 8. Вигляд b-структури – складчастий лист, де R-радикали амінокислот Інша різновидність b-структури має назву паралельного складчастого шару. Утворюється вона в тому випадку, коли N -кінці поліпептидних ланцюгів напрямлені в один бік.
Рис. 9. Схематичне зображення b-структур: а -паралельні ланцюги, б -антипаралельні ланцюги Для b-конформації характерні міжланцюгові водневі зв'язки між пептидними групами сусідніх поліпептидних ланцюгів. Для того, щоб сусідні ділянки білкової послідовності розташувалися в орієнтації, котра сприяє утворенню міжланцюгових водневих зв'язків, між ними має знаходиться неструктурована ділянка, здатна утворити b-вигин. Якщо складчастий ланцюг вигинається, робить поворот назад і йде вздовж самого себе назад – це антипаралельна b-конформація. Місце повороту – b-вигин. (рис. 10).
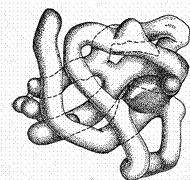
Рис. 10. Антипаралельна b-структура білка та b-вигин b-Структура характерна для білків опорних тканин – колагену (білок сухожилля, шкіри), фіброїну (білок шовку), окремих видів кератину (білок шерсті, волосся) та ін. У деяких білках можливі переходи від a-спіралі до Отже,a-спіраль та b-структура становлять другий порядок, або рівень, структурної організації молекули білка, тобто вторинну структура, в стабілізації якої важливу роль відіграють водневі зв'язки. Яку саме конформацію приймають ділянки поліпептидного ланцюга Третинна структура білка – це просторова конформація поліпептиду, що має вторинну структуру, і обумовлена взаємодіями між радикалами. Усі біологічні властивості білків пов'язані з цілісністю їх третинної структури, яку прийнято називати нативною конформацією. Будь-які порушення нативної конформації призводять до часткової або повної втрати біологічної функції. Структура біополімерів найтіснішим чином пов'язана з їхньою біологічною функцією. Для того, щоб виконувати «будівельні» опорні функції (волосяний покрив, зміцнення стінок клітини, цитоскелет клітини і т.п.) білок повинен мати витягнуту лінійну структуру і не розчинятися у воді. Такі білки називаються фібрилярними. Структура фібрилярних білків (колаген, кератин волосся, еластин судин). Що б володіти витягнутою нитковидною структурою, білок має бути високо структурований, тобто мати вторинну структуру (a-спіральної або Структура глобулярних білків. Крім структурних функцій білки виконують інші функції. Серед них, одна з головних – каталіз хімічних реакцій, що протікають у клітині. Для здійснення каталітичної (ферментативної) функції білок має зв'язуватися з тривимірним субстратом, тобто місце зв'язування, назване активним центром, повинне бути тривимірним. А це можливо тільки за умови, що сам білок має тривимірну третинну (глобулярну) структуру. Крім того, глобулярна структура забезпечує розчинність білків у воді (більшість реакцій протікає в цитоплазмі клітини). Гідрофобні радикали амінокислотних залишків можуть бути заховані у середину глобули, а у воду експоновані тобто направлені гідрофільні залишки. Для забезпечення компактної тривимірної структури білок повинен мати елементи вторинної структури невеликої довжини, з'єднаних не структурованими ділянками поліпептидного ланцюга. Ці ділянки дозволяють білку скручуватися у компактну глобулу.
Рис. 11. Глобулярна третинна структура білка міоглобіну. Третинна структура цілком задається первинною. (рис. 11). Визначальними є гідрофобні взаємодії, водневі, дисульфідні та йонні зв¢язки. Гідрофобне ядро існує в більшості білків, гідрофільні групи залишків амінокислот розміщені назовні. Третинна структура є складною, унікальною і специфічною для кожної молекули білка. Четвертинна структура білка – це агрегація двох або більшого числа поліпептидних ланцюгів, що мають третинну структуру, в олігомерну функціонально значущу композицію. Тобто утворюється єдина у структурному та функціональному відношенні одиниця. Білки, що складаються з декількох структурних одиниць називаються олігомерними. Окремі структурні одиниці олігомерного білка називають субодиницями. Взаємне розташування субодиниць, тобто спосіб їхнього спільного укладання й упакування з утворенням нативної конформації олігомерного білка і називають четвертинною структурою. Олігомерний білок може містити дві, три, чотири або більше субодиниць. Іноді зустрічаються олігомери, що складаються з 12 субодиниць. Субодиниці четвертинної структури можуть бути як однаковими, так і різними, розташовуватись у четвертинній структурі як симетрично, так і асиметрично. Зв'язки, що утворюють і підтримують четвертинну структуру, ті ж самі, що і при утворенні третинної структури, крім гідрофобних. Четвертинну структуру має близько 5% білків, у тому числі гемоглобін, імуноглобулін, інсулін. Майже всі ДНК- і РНК-полімерази мають четвертинну структуру. 3.2.4. Класифікація білків · Прості білки – побудовані тільки із залишків амінокислот, при гідролізі дають вільні амінокислоти. 1)Протаміни. Сильно лужні білки з невеликою молекулярною масою. Складаються на 60 – 85% з аргініну. Добре розчинні у воді. 2) Гістони.Білки з основними властивостями Складаються на 20 – 30% з лізину та аргініну. Розташовані переважно у ядрі, стабілізують ДНК. 3)Проламіни. Розчинні у 60 – 80% водному розчині етанолу, усі інші прості білки за таких умов випадають в осад. Містять в своєму складі 20 – 25% глутамінової кислоти та 10 – 15% проліну. 4)Глютеліни. Розчинні в лужних розчинах. Складаються в переважній більшості з глютаміну та лізину. Проламіни та глютеліни це в основному білки рослинного походження, складають основну масу клейковини зерен злаків. 5) Альбуміни. Розчинні у воді та розчинах солей. Молекулярна маса: альбумінів – 40000 – 70000. Альбуміни та глобуліни – це глобулярні білки, які є основними білками плазми крові. Альбуміни складають 60 - 65% загального білка плазми крові. 6) Глобуліни. Мають більшу молекулярну масу і розчинність у солевих розчинах низької концентрації. Молекулярна маса понад 150000. Найбільше цих білків у сироватці (плазмі) крові, молока, яєчному білку, м’язах. Глобуліни виробляються печінкою та імунною системою. · Складні білки – це двохкомпонентні білки, які складаються із простого білка та небілкової компоненти, яка називається простетичною групою (prosteto – приєдную) Класифікація складних білків – за природою простетичної групи: Хромопротеїни. Побудовані з простого білка і забарвленої простетичної групи (пігменту). Розрізняють гемопротеїни (містять гем), магнійпорфірини і флавопротеїни (містять похідні ізоалоксазину). Хромопротеїни мають ряд унікальних біологічних функцій: вони беруть участь в таких фундаментальних процесах життєдіяльності, як фотосинтез, клітинне дихання і транспорт кисню і диоксиду вуглецю у межах цілого організму, окисно-відновні реакції, сприйняття світла і кольору та інші. Металопротеїни. До складу простетичної групи входять залізо, мідь, кобальт, цинк та інші елементи. До металопротеїдів належить велика група ферментів. Нуклеопротеїни. Це складні білки, простетичною групою яких є нуклеїнові кислоти. Залежно від природи нуклеїнової кислоти нуклеопротеїди поділяються на рибонуклеопротеїди (РНП), якщо до їх складу входить рибонуклеїнова кислота – РНК, а якщо дезоксирибонуклеїнова кислота – ДНК. Ліпопротеїни. У складі простетичної групи містять ліпіди і їх похідні. Найчастіше простетичною групою є нейтральні жири – тригліцериди. Входять до складу біологічних мембран, крові, лімфи, нервових тканин та інше. В комплексі з білками ліпіди набувають нових властивостей: вони здатні розчинятися у воді і втрачають здатність розчинятися у органічних розчинниках. Фосфопротеїни. Мають в своєму складі залишок фосфатної кислоти. Ці білки входять до складу білків яєць (альбумін), молока (казеїн). Глікопротеїни. Містять залишки вуглеводів. Входять до складу крові, молока, яєць, ферментів, гормонів. До цієї групи належать імуноглобуліни, деякі гормони: гонадотропні, тиреотропін, кортикотропін, тиреоглобулін, інгібітор розмноження вірусів – інтерферон. 3.2.5. Якісне та кількісне визначення білків Присутність білків у харчових об'єктах встановлюється за допомогою якісних реакцій, які умовно поділяють на дві групи: а) кольорові реакції; б) реакції осадження. Серед першої групи розрізняють універсальні реакції (біуретова реакція Піотровського на пептидні зв'язки та нінгідринова на α-амінокислоти) та специфічні, зумовлені реакціями функціональних груп певних амінокислот присутніх в білках. Так, ксантопротеїнова реакція свідчить про наявність в білках залишків ароматичних амінокислот, реакція Паулі – гістидину і тирозину, Адамкевича і Вуазене – триптофану, нітропрусидна – цистеїну, а реакція Сакагучи – аргініну. За результатами специфічних реакцій орієнтовно можна судити про харчову цінність білків. Біуретова реакція. У лужному середовищі в присутності іонів Купруму (ІІ) розчини білків і пептидів набувають фіолетового забарвлення з червоним або синім відтінком залежно від кількості пептидних зв'язків:
Біуретовий мідний комплекс фіолетового кольору. Нінгідринова реакція на a-аміногрупу.
Амінокислота Нінгідрин Відновлений нінгідрин
Сполука з синьо-фіолетовим забарвленням Ксантопротеїнова реакція. Ця реакція характерна для амінокислот з ароматичним кільцем (реакція Мульдера). В результаті нітрування бензенового кільця утворюється жовтий осад, який в надлишку лугу переходить в помаранчеве забарвлення.
Тирозин Динітротирозин Хіноїдна форма (жовто-оранжеве забарвлення) У другій групі реакцій білки осаджують в ізоелектричній точці, дією солей, органічних розчинників, концентрованих кислот, лугів, йонів важких металів, температури. Ізоелектричною точкою (ІЕТ) білка називається концентрація йонів гідрогену, при якій білок знаходиться в ізоелектричному стані (сумарна кількість негативних і позитивних зарядів однакова),тобто незаряджений. ІЕТ характеризує хімічну природу білка. Для кожного білка існує своя ІЕТ. При рН, близькому до ІЕТ, розчинність, набухання, в'язкість білка стають найменшими, а осадження, аглютинація та комплексоутворення – найлегшими. Таким чином, в ІЕТ білок нестабільний і легко випадає в осад, особливо в присутності водовіднімних речовин (спирту, ацетону та ін.). Білки в розчиненому стані вкрай нестійкі, тому при додаванні органічних розчинників (спирт, ацетон), концентрованих розчинів нейтральних солей лужних металів гідратна оболонка руйнується і вони випадають в осад. Порушення гідратної оболонки навколо молекул білка під дією солей лужних та лужноземельних металів, що призводить до випадіння білків в осад називається висолюванням. Як правило після видалення солі білок не втрачає здатності розчинятися у воді. Різні білки випадають в осад при різних концентраціях нейтральних розчинів сульфату амонію. На величину висолювання білків впливає природа солі, її концентрація, рН розчину і температура. Білки під впливом фізичних (температури, ультразвуку, іонізуючої радіації та інших), хімічних (мінеральних і органічних кислот, лугів, органічних розчинників, важких металів, алкалоїдів тощо) факторів зазнають глибоких змін, пов'язаних з порушенням четвертинної, третинної і вторинної структури, що призводить до зміни фізико-хімічних і біологічних властивостей білка, тобто до денатурації (втрати нативності). При денатурації білка відбувається розрив «цементуючих» білкову молекулу вторинних зв'язків (водневих, дисульфідних, електростатичних, ефірних, вандервальсових та ін.). Це призводить до зміни просторової структури і зменшує гідрофільні властивості білка. Він стає більш гідрофобним, втрачає здатність розчинятись у звичайних для нього розчинниках і втрачає свої біологічні функції. Глибока денатурація є незворотною на відміну від взаємно-зворотної (ренатурації), при якій зміни структури білка бувають неглибокими і білок за деяких умов може знову набувати своїх нативних властивостей. Наприклад, при осадженні білків органічними розчинниками – спиртом або ацетоном (при низькій температурі), з подальшим видаленням осаджувача. Для кількісного визначення білків найчастіше застосовують фотоколориметричні і спектрофотометричні методи, у деяких випадках використовують визначення білка за вмістом загального нітрогену (азотометрія), а також фотонефелометричні методи. 3.2.6. Функціональні властивості білків Під функціональними властивостями розуміють фізико-хімічні властивості, які визначають зміни білків при переробці у харчові продукти та забезпечують певну структуру, технологічні і споживчі властивості. До найбільш важливих характеристик білків належать: водо- і жирозв‘язуюча здатність, структуроутворююча (піноутворююча, драглеутворююча) здатність. Здатність білків зв'язувати на своїй контактній поверхні воду – одна із характерних фізико-хімічних властивостей, що відіграє важливу роль у технології харчових продуктів. З гідрофільністю пов'язані такі процеси, як набухання і розчинність білків, їх осадження і денатурація при дії жорстких фізико-хімічних факторів середовища. Гідрофільність являє собою наслідок дії електростатичних сил притягування, що розвиваються між йоногенними і полярними групами білкової глобули і диполями води. Набухання. Внаслідок великої різниці у швидкості дифузії молекул білків і низькомолекулярного розчинника (вода) на першій стадії процесу розчинення білка молекули розчинника проникають у простір між його полімерними ланцюгами. Відбувається збільшення об'єму молекули. Це явище називають набуханням, яке переходить у розчинення (необмежене набухання) у тому випадку, коли між поліпептидними ланцюгами відсутні поперечні хімічні зв'язки. Це явище характерне для глобулярних білків. У поліпептидах, що утворюють сітчасту структуру, розчинення неможливе, і процес закінчується набуханням (обмежене набухання). Такі перетворення характерні для деяких фібрилярних білків. Кількісною характеристикою обмеженого набухання є ступінь набухання αm яку визначають гравіметричним способом: зважують зразок до і після набухання і розраховують за формулою: αm= т-т0/то де т – маса набухлого зразка; т о – наважка вихідного зразка. Ступінь набухання також визначають об'ємним методом, вимірюючи об'єм зразка до і після набухання: αv =V-V0/ V0 де V – об'єм набухлого зразка;V0 – об'єм вихідного зразка. Ступінь набухання залежить від різних факторів, у тому числі від природи розчинника. При контакті з водою сухий білок набухає, молекули води проникають у білкову масу і в результаті гідратації молекули білка роз'єднуються. Важливу роль тут відіграють не тільки електростатичні сили, але і сили осмосу. Набухлий білок можна вважати розчином води у білку: енергія набухання велика. Подальше поглинання води призводить до розчинення білка. Розчинення пов'язано з хімічною структурою білка. Оскільки амінокислотний склад білків різний, розчинність білків коливається у широких межах і буде визначатись співвідношенням гідрофільних (полярних) і гідрофобних (неполярних) груп,специфікою укладання їх у трьохвимірну структуру; чим більше гідрофільних груп на поверхні білкової молекули, тим вища її гідрофільність і тим вища розчинність білка; поверхневі гідрофобні групи зменшують розчинність. Гло   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры...  ЧТО И КАК ПИСАЛИ О МОДЕ В ЖУРНАЛАХ НАЧАЛА XX ВЕКА Первый номер журнала «Аполлон» за 1909 г. начинался, по сути, с программного заявления редакции журнала...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
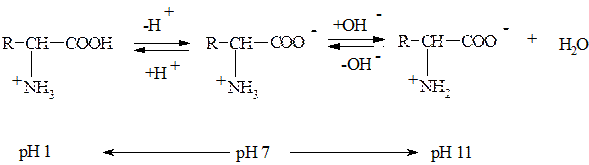


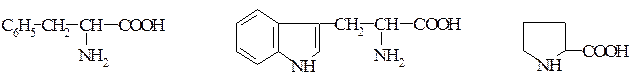
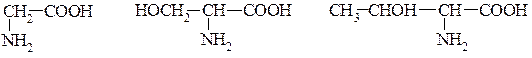
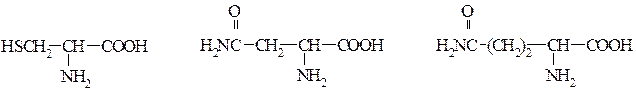
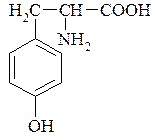

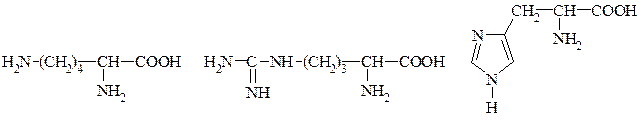 Лізин Аргінін Гістидин
Лізин Аргінін Гістидин
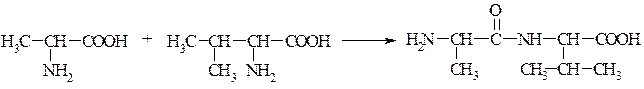 АланінВалін Дипептид (аланіл-валін)
АланінВалін Дипептид (аланіл-валін)