|
ГЛАВА 3. МЕЗОЗОЙСКАЯ ЭРА - ВЕК РЕПТИЛИЙ
После завершения в середине пермского периода оледенения Гондваны началось потепление, которое продолжалось на протяжении мезозойской эры, в целом более однообразной в климатическом отношении, чем другие эры фанерозоя. В мезозое господствовали теплые климаты с относительно слабо выраженной климатической зональностью. Следов оледенения какого-либо континента в мезозое не обнаружено. Южный полюс оставался в океане, а Северный перемещался от восточной оконечности Сибири к Аляске. При полном отсутствии оледенений температуры воздуха и воды в океане были, вероятно, значительно выше современных: на экваторе на 3-5 °С, в средних широтах на 10 С, а в полярных - на 20-40 °С.
В то же время происходили существенные изменения рельефа Земли и положения континентов, которые привели к формированию в общих чертах существующих ныне континентов и океанов. В начале триаса сохранялись относительно высокие материки. Затем началась морская трансгрессия с образованием мелких краевых морей Тихого океана, прерывавшаяся в начале и в конце юрского периода Древне- и Позднекиммерийскими фазами горообразования. В меловом периоде новая морская трансгрессия привела к затоплению значительных территорий Европы и Северной Африки краевыми теплыми морями древнего океана Тетис.
По представлениям теории тектоники плит, общей тенденцией в мезозое был распад суперконтинента Пангеи и расхождения континентальных плит (рис. 60). В триасе этот процесс только начался. Лавразия сместилась несколько к северу. При этом происходило раскрытие океана Тетис, отделявшего Лавразию от Гондваны. Затем Лавразия раскололась на Северную Америку и Евразию, между которыми, начиная с раннеюрского времени, началось формирование северной части Атлантического океана. Вероятно, уже к концу юрского периода начался раскол Гондваны и, соответственно, возникновение Индийского океана и южной части Атлантики, отделивших Африку от Австралии и Антарктиды на востоке и юге и от Южной Америки на западе. Соединение северной и южной частей Атлантики произошло уже в позднемеловое время.
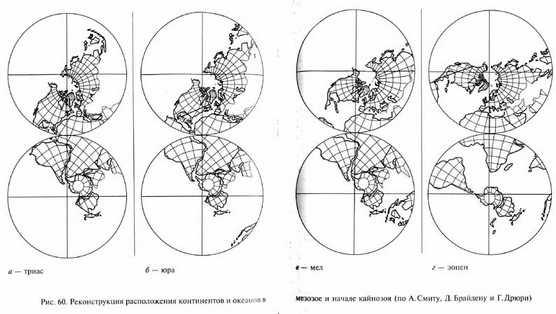
В меловом периоде расхождение континентов, которые в конце палеозоя входили в состав единой Пангеи, значительно прогрессировало. Продолжали расширяться Атлантический и Индийский океаны. При движении американских плит к западу на их западных окраинах возникли мощные зоны горной складчатости: в юрском периоде - Сьерра-Невада, в меловом - Анды и Скалистые горы (Позднекиммерийская и Ларамийская фазы горообразования). Индостан и Мадагаскар обособились от Африки, Индостанская плита начала смещение к северо-востоку. Сложные взаимные смещения Африки и Европы привели к постепенному закрытию океана Тетис. Из материков, входивших в состав Гондваны, в конце мезозоя остались соединенными лишь Антарктида и Австралия. Зато материки, составлявшие Лавразию и разделившиеся в начале мезозоя, к концу мела вновь соединились, правда противоположными концами: Азия и Северная Америка сошлись в области Чукотки и Аляски. Так возникла Берингия, сухопутный мост, связавший Америку и Европу на месте современного Берингова пролива.
Обновление флоры и фауны в триасе
Пермское оледенение и последовавшее после его завершения глобальное потепление климата привели к значительному изменению общего облика флоры. Вымерли древовидные плаунооб-разные, столь характерные для лесов второй половины палеозоя. Исчезли также семенные папоротники (птеридоспермы) и кордаиты. Их место занимают разнообразные представители других групп голосеменных растений: саговниковых (Cycadophyta), гинк-говых (Ginkgoales) и хвойных (Coniferales) (см. рис. 68, изображающий ландшафт юрского периода, где представлены указанные голосеменные, появившиеся уже в триасе). Особенно характерны для наземной флоры первой половины мезозоя были саговниковые: древовидные пальмообразные, с высокими колоннообразными стволами, увенчанными пучком перистых листьев (Williamsonia), или приземистые, с бочкообразным или клубневидным коротким и толстым стволом (Cycadeoidea). Саговниковые, хвойные и гинкговые формировали общий облик лесов в первой половине мезозойской эры.

Рубеж между пермским и триасовым периодами был отмечен также существенными изменениями в фауне наземных позвоночных. В пермское время преобладающими группами пресмыкающихся были зверообразные рептилии и котилозавры, тогда как группы с диапсидным черепом (лепидозавры и архозавры), объединяемые под названием "завропсиды", оставались сравнительно Немногочисленными. Это соотношение резко изменилось к началу мезозоя. Из числа котилозавров в триас перешли лишь проколофоны (вымирающие к концу этого периода), а из зверообразных - немногие группы дицинодонтов и высших териодонтов. Зато обилие и разнообразие диапсидных рептилий неуклонно возрас-Тает, и во второй половине триаса они становятся доминирую-Щими. При этом нужно отметить, что высшие териодонты обладали рядом прогрессивных признаков, отсутствовавших у завропсид (например, были способны к пережевыванию пищи, значительно повышающему степень ее усвоения организмом; вероятно, они имели теплоизолирующий волосяной покров и т.д.). Победа в борьбе за существование была достигнута завропсидами, так сказать, вопреки прогрессивным чертам организации териодонтов и, очевидно, основывалась на каких-то важных преимуществах первых.
П.Робинсон предложила гипотезу, связывающую изменения фауны рептилий в триасе с особенностями физиологии завропсид и зверообразных пресмыкающихся, о которых с известной долей вероятности можно судить по соответствующим характеристикам современных потомков тех и других. Современные завропсиды (в широком смысле этот термин используется для объединения всех современных групп этого класса и птиц) характеризуются наличием целого комплекса приспособлений к жизни в условиях жаркого засушливого климата. Завропсиды могут выдержать значительно более высокие температуры тела, чем потомки зверообразных рептилий - млекопитающие (многие ящерицы - до 44 °С, птицы - до 43 °С, млекопитающие - лишь до 39 °С; для Млекопитающих смертельно повышение температуры тела до 43 °С, Для птиц - до 46-47 °С). При этом рептилии используют энергию солнечного облучения для повышения температуры своего тела До оптимального уровня (гелиотермия); в связи с этим для них сохраняет важное значение "теменной глаз", регулирующий различные функции организма в зависимости от интенсивности падающего света.
Конечным продуктом белкового обмена у завропсид является мочевая кислота, тогда как у млекопитающих - мочевина. Мочевая кислота может образовывать перенасыщенные растворы, поэтому для ее выведения из организма требуется примерно в 10 раз меньше воды, чем для выведения мочевины. Очевидно, у завропсид выделительная система гораздо лучше экономит воду для организма, чем у млекопитающих.
Логично предположить, что организация завропсид складывалась под контролем отбора, приспосабливавшего этих рептилий к условиям засушливого и жаркого климата, тогда как зверообразные рептилии сформировались в местообитаниях с более влажным и прохладным климатом. Такие местообитания, вероятно, были широко распространены на континентах, входивших в состав Гондваны, в эпоху великого палеозойского оледенения. К концу пермского периода и в триасе во многих регионах климат становится все более засушливым (или, по крайней мере, характеризовался значительными сезонными засухами). П.Робинсон связывает это с изменениями морских течений и преобладающих направлений циркуляции атмосферных потоков в результате дрейфа континентов, входивших в состав Лавразии и Гондваны. Засушливость климата должна была дать важные преимущества в борьбе за существование тем группам наземных животных, которые обладали более высокой устойчивостью к действию высоких температур, были способны лучше экономить воду и с помощью специальных форм поведения могли использовать энергию солнечного облучения для достижения оптимальной температуры своего тела. Поскольку по всем этим показателям завропсиды превосходили зверообразных рептилий, последние были оттеснены в еще сохранившиеся более прохладные и влажные местообитания. В неблагоприятных для них условиях, при жесткой конкуренции со стороны завропсид, численность зверообразных рептилий значительно упала, и большинство их групп вымерло.
В немногих сохранившихся в триасе филогенетических стволах высших териодонтов продолжались изменения, наметившиеся еще среди пермских терапсид. Общее направление этих эволюционных изменений обозначают иногда термином "маммализация", т. е. развитие комплекса признаков, характерных для высших амниот -млекопитающих. Здесь мы вновь встречаемся с параллельной эволюцией: сходные черты строения независимо возникали в разных филетических линиях териодонтов. Среди этих признаков было постепенное увеличение размеров полушарий переднего мозга, приобретение мягких губ (что делало возможным сосание) и волосяного покрова. По мнению Л.П.Татаринова, зачаточные волоски у териодонтов имели осязательную функцию, располагаясь вблизи рта (как специализированные осязательные волосы - вибриссы у млекопитающих). Однако в среднетриасовых отложениях были обнаружены ископаемые следы цинодонта (Cynodontia - "собакозубые", одна из групп высших териодонтов) с отпечатками волос. Поэтому имеются основания считать, что у териодонтов волосяной покров был развит уже на всем теле, как у млекопитающих, и, вероятно, имел ту же основную функцию, как и у последних, т. е. служил для теплоизоляции (это понятно, если принять гипотезу о возникновении и развитии зверообразных рептилий в прохладном климате).

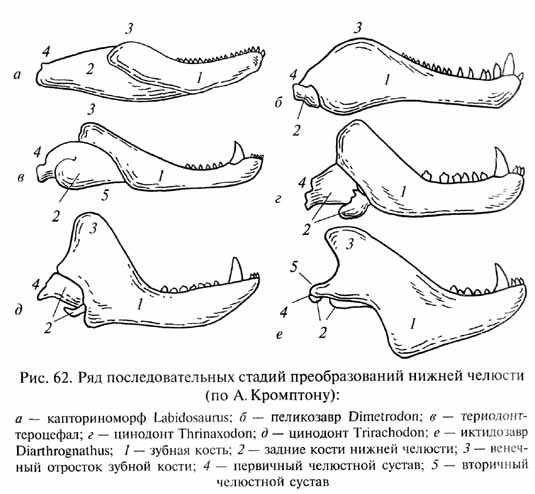
У цинодонтов (рис. 61) продолжалось совершенствование механизма пережевывания пищи с развитием окклюзии (смыкания коронок верхне- и нижнечелюстных зубов в щечной области) и продольных и поперечных движений нижней челюсти. Для обеспечения этих движений произошли сложные перестройки челюстных мышц, сыгравшие важную роль в дальнейшей эволюции челюстного аппарата. Механически наиболее благоприятное расположение челюстных мышц было достигнуто при разрастании зубной кости нижней челюсти назад и вверх с развитием высокого венечного отростка, к которому прикреплялась височная мышца (рис. 62). При этом задние кости нижней челюсти подверглись редукции. Гипертрофия задней части зубной кости Привела к возникновению ее контакта (при движениях нижней Челюсти) с чешуйчатой костью черепа. Между двумя этими костями возникла суставоподобная связь, которая оказалась механически более эффективной опорой и осью вращения для нижней челюсти, чем первичный челюстной сустав (между квадратной Костью верхней и сочленовной костью нижней челюсти). Преобразования нижней челюсти оказались преадаптивными для формирования новой опоры нижней челюсти - вторичного челюстного сустава.
Вторичный челюстной сустав, присущий среди всех позвоночных только млекопитающим, стал поэтому важнейшим диагностическим признаком последних. Редуцированные кости первичного челюстного сустава, освободившиеся от своей прежней функции, вошли в состав цепи косточек среднего уха в качестве двух наружных ее элементов - наковаленки и молоточка, что стало еще одной характерной особенностью млекопитающих, обеспечившей более тонкую слуховую чувствительность в области высоких звуковых частот.

Наиболее древние ископаемые остатки млекопитающих или зверей (Mammalia) известны из верхнетриасовых отложений. Это были небольшие хищные животные (рис. 63), питавшиеся различными мелкими позвоночными и беспозвоночными. Вероятно, они предпочитали прохладные и влажные местообитания и вели преимущественно ночной образ жизни. Скорее всего, у них уже существовало выкармливание детенышей молоком, поскольку оно присуще всем современным группам млекопитающих, а расхождение филогенетических стволов, ведущих, с одной стороны, к однопроходным (Monotremata), с другой - к сумчатым (Marsupialia) и плацентарным (Placentalia), произошло, возможно, уже в позднем триасе. Млечные железы возникли у древних млекопитающих из видоизмененных потовых желез. Возможно, первоначальной функцией млечных желез было не столько выкармливание, сколько "выпаивание" детенышей, т. е. снабжение их необходимой влагой и солями. Дефицит влаги был особенно опасен для новорожденных детенышей млекопитающих в условиях засушливого климата. Судя по способу развития однопроходных, наиболее примитивных среди современных млекопитающих, древнейшие звери еще были яйцекладущими животными, которые высиживали свою кладку или вынашивали яйца в специальной сумке на животе.
В конце триаса млекопитающие уже были довольно разнообразны и принадлежали к нескольким обособленным группам: три-конодонтам (Triconodontia), симметродонтам (Symmetrodontia) и харамиидам (Haramiyidae), различавшимся по строению зубов и, вероятно, по способам питания и пережевывания пищи.
Несмотря на ряд прогрессивных морфофизиологических особенностей (некоторое увеличение головного мозга, особенно его больших полушарий; забота о потомстве и выкармливание его молоком; пережевывание пищи; волосяной покров, расположение конечностей под туловищем, обеспечивающее более совершенную локомоцию), млекопитающие в мезозое не достигли больших успехов в конкуренции с рептилиями. Они оставались относительно малочисленными мелкими животными и, вероятно, вели ночной или сумеречный образ жизни, характерный для большинства зверей и ныне. Возможно, именно несовершенство систем терморегуляции и выделения, приспособленных к условиям относительно низких температур и высокой влажности, снижали для мезозойских млекопитающих шансы на успех в конкуренции с завропсидами. Хотя мезозойские млекопитающие уже обладали теплоизолирующим шерстным покровом и, вероятно, могли усилить теплоотдачу за счет потоотделения, у них не было еще совершенной интеграции разных терморегуляционных механизмов в единую систему. Об этом можно судить по состоянию современных однопроходных, у которых еще нет настоящей гомойотермии, и температура тела колеблется в пределах 26-34 °С.
В триасе начался подлинный расцвет рептилий. Особенно многочисленными становятся лепидозавры и архозавры. На ранних этапах их эволюционной истории эти два филогенетических ствола оставались еще очень близкими друг к другу, поэтому нередко трудно с достаточной уверенностью дифференцировать примитивных лепидозавров и архозавров. Таковы, например, ринхозавры (Rhynchosauria), которых одни авторы сближают с клювоголовы-ми среди лепидозавров, а другие относят к архозаврам. Ринхозавры получили широкое распространение в триасовое время. Это были крупные животные (1,5-5 м) с широким черепом, мошной жевательной мускулатурой и многорядными зубами на верхнечелюстных и зубных костях. Ринхозавры были растительноядными и, вероятно, конкурировали с дицинодонтами.
Настоящие клювоголовые (ринхоцефалы), родственные современной гаттерии, известны начиная с позднего триаса. В среднем триасе уже существовали ящерицы (группа эолацертилий), которые вскоре становятся очень разнообразными. Например, средне-триасовый танистрофей (Tanystropheus) обладал гипертрофированной шеей, длина которой превышала длину туловища (рис. 64, б)', общие размеры животного достигали 6 м. Возможно, эти рептилии, которых некоторые ученые также считают не ящерицами, а архозаврами, вели водный образ жизни, используя свою феноменальную шею при ловле рыб и головоногих моллюсков, как более поздние плезиозавры. Среди позднетриасовых эолацертилий имелись формы (Kuehneosaurus, Icarosaurus), приспособившиеся к планирующему полету: на удлиненных ребрах, выступавших по бокам туловища, была натянута летательная перепонка (рис. 64, а).

Возможно, вымирание последних котилозавров - проколофонов - в конце триаса было связано с их неудачей в конкуренции с экологически близкими, но более совершенными ящерицами.
Многочисленны и разнообразны в триасе были и представители архозавров. Корневой группой этого мощного филогенетического ствола являются псевдозухии, или текодонты (Pseudosi или Thecodontia), - преимущественно небольшие животные, примитивные представители которых внешне несколько напоминали ящериц. Но у псевдозухии череп сохранял типичное диапсидное строение, а зубы сидели в ячейках (текодонтный тип), а не прикреплялись к краю челюсти, как у ящериц. Для псевдозухии характерна тенденция к удлинению задних конечностей, связанная с переходом к быстрому бегу на двух ногах (рис. 65). Нужно отметить, что и ящерицы при максимальной скорости бегут лишь на двух задних конечностях, приподняв переднюю часть тела и длинный хвост над субстратом. Такой способ убыстрения бега (связанный с сохранением примитивной постановки конечностей, с горизонтальным положением плеча и бедра) был, видимо, характерен и для псевдозухии, но у последних получил более значительное развитие. Среди их потомков появились настоящие двуногие формы. Большинство псевдозухии были хищниками мелких и средних размеров (самые крупные достигали 3,5 м в длину). Некоторые из них стали быстро бегающими наземными животными, другие приспособились к лазанию по деревьям, были также полуводные формы - фитозавры (Phytosauria), имевшие внешнее сходство с крокодилами. В позднем триасе от псевдозухии возникли и настоящие крокодилы (Crocodylia), оказавшиеся лучше приспособленными к роли амфибиотических хищников и быстро вытеснившие фитозавров.

И фитозавры, и крокодилы конкурировали в водоемах с последними стегоцефалами - лабиринтодонтами, представленными в триасе очень крупными формами с огромным уплощенным черепом, достигавшим у Mastodonsauras 125 см длины. Однако и эти гиганты не выдержали конкуренции с водными рептилиями. Разнообразие и численность стегоцефалов в первой половине мезозоя неуклонно сокращались, и последние стегоцефалы (плагиозавры) вымерли в раннемеловое время.
В триасе появились также черепахи (Chelonia). Костно-роговой двуслойный панцирь черепах оказался замечательной защитной адаптацией, позволившей этим рептилиям сохранить многие примитивные черты организации, соответствующие уровню котилозавров. Происхождение черепах до сих пор остается дискуссионным. Древнейший известный их представитель, позднетриасовый Proganochelys, уже имел основные черты строения черепах. Разные исследователи видели предков черепах в различных группах котилозавров (от капториноморфов до парейазавров). М.Ф.Ивахненко, основываясь на некоторых особенностях строения черепа, высказал гипотезу о происхождении черепах от особого филогенетического ствола, названного Е.Олсоном парарептилиями и эволюционировавшего от уровня стегоцефалов независимо от других пресмыкающихся. Однако эта гипотеза требует допущения, Что все основные особенности амниот, рассмотренные нами выше (скелет, кожные покровы, дыхательная, кровеносная и выделительная системы, а также амниотический онтогенез), развивались независимо и параллельно у настоящих рептилий и парарептилий. Это представляется менее вероятным, чем параллельное (и неоднократное) формирование сходных особенностей черепа в разных линиях эволюции рептилий. Решение проблемы происхождения черепах, очевидно, ждет новых палеонтологических данных.
Жизнь в мезозойских морях
Теплые моря, заливы и лагуны в мезозое были богаты жизнью. После пермского вымирания разнообразие морских организмов в триасе вновь возрастает. По-видимому, в мезозое возник новый тип водорослей - диатомовые (Diatomeae). Во всяком случае, их ископаемые остатки достоверно известны, начиная лишь с юрских отложений.
В донной фауне опять обильны мшанки, многочисленны морские ежи и морские звезды. Постепенно возрастает численность и разнообразие брюхоногих моллюсков. Среди двустворчатых моллюсков выделяется мезозойская группа рудистов (Rudistae), прикреплявшихся к субстрату одной из створок своей асимметричной раковины, достигавшей у некоторых видов 1,5 м. В триасе начинается бурный расцвет аммонитов и белемнитов. Мезозойские головоногие были очень разнообразны и по форме раковины, и по размерам, варьировавшим у аммонитов от нескольких миллиметров до 3 м. Белемниты, подобно современным кальмарам, стремительно проносились в толще воды, охотясь за мелкими рыбами, а аммониты медленно плавали, регулируя глубину погружения с помощью раковины - гидростатического аппарата.
Постепенно обновилась ихтиофауна. В юрское время сформировались современные группы пластинчатожаберных хрящевых рыб (Elasmobranchia): акулы (Selachoidea) и скаты (Batoidea). Среди костных рыб появились новые группы лучеперых: костные ганои-ды (Holostei) и костистые рыбы (Teleostei). Первые пережили свой расцвет в юре, вторые возникли в юрское время, а начиная с мела стали доминирующей группой (к костистым принадлежит 95% всех современных видов рыб). Продолжали существовать в мезозойских морях кистеперые - целаканты.
Хрящевые рыбы (Chondrichthyes) полностью утратили костный скелет, характерный для всех древнейших палеозойских рыб. Это было, вероятно, связано с необходимостью освободиться от тяжелого наружного костного панциря при активизации образа жизни. Костные рыбы (Osteichthyes) также решали проблему облегчения костного панциря, но не путем его утраты вместе с внутренним костным скелетом, а посредством постепенных и более длительных преобразований, которые привели к формирований гибкого, легкого и прочного чешуйного покрова. Можно сказать, что в отношении преобразований наружного и внутреннего скелета хрящевые рыбы в сравнении с костными пошли по инадаптивному пути. В конкуренции с костными рыбами хрящевые рыбы компенсировали несовершенство своего скелета увеличением размеров тела и рядом прогрессивных изменений организации: совершенствование головного мозга, внутреннее осеменение, откладка крупных яиц, защищенных прочными оболочками, снабженных большим количеством желтка и обеспечивающих длительное развитие зародыша.
Всякая группа организмов, сумевшая в ходе эволюционных преобразований достичь нового уровня организации, под давлением естественного отбора начинает экологическую экспансию, осваивая все доступные местообитания, способы питания и т.п. При этом закономерно происходит адаптивная радиация - расхождение признаков у потомков общей предковой группы в процессе приспособительной эволюции, ведущее к возникновению разнообразных видов. Разнообразие развивающихся при этом приспособлений и возникающих жизненных форм зависит от совершенства и пластичности организации данной группы. Несомненно, одной из самых поразительных в этом отношении групп являются рептилии, которые в мезозое освоили практически все возможные типы местообитаний и дали фантастическое разнообразие жизненных форм, не превзойденное ни одним другим классом позвоночных животных.
Триас ознаменовался успешным освоением рептилиями не только пресных водоемов (танистрофеи, фитозавры и крокодилы), но и морей. Морские рептилии получили в мезозое широкое распространение и были чрезвычайно разнообразны: одни из них жили на мелководье в прибрежной зоне и, вероятно, сохраняли некоторую связь с сушей, другие же стали обитателями открытого океана и никогда не выходили на берег. Среди первых наиболее известны завроптеригии (Sauroptcrygia). Это были животные довольно странного облика (рис. 66): массивное широкое туловище, имевшее, однако, обтекаемую форму, впереди продолжал ось длинной и тонкой шеей (у некоторых форм она была длиннее туловища), а сзади - длинным сжатым с боков веслообразным хвостом, несущим сверху небольшой хвостовой плавник. Передние и задние конечности у более специализированных к жизни в море плезиозавров (Plesiosauria) имели строение широких и мощных ластов, которые были основными движителями в воде. Хвостовой Плавник был невелик и мог помогать при плавании с малой скоростью, а в основном он, вероятно, служил рулем. Ласты могли служить опорой при выходе плезиозавров на прибрежные отмели Для откладки яиц. как это делают современные морские черепахи. Однако даже такое кратковременное возвращение на сушу для Некоторых плезиозавров было очень затруднено их огромными размерами: ряд юрских и меловых видов достигал 12-16 м (плиозавры, элазмозавры).

Причудливый облик плезиозавров и упоминавшихся выше та-нистрофеев имеет современную аналогию, хотя и в миниатюре: у морских змей-ластохвостов (Hydrophis) также имеется утолщенное туловище, плоский хвост и длинная тонкая шея (в 4-5 раз более тонкая, чем туловище). Такие пропорции тела помогают водному хищнику использовать массивное и широкое туловище как своего рода опору в воде на плаву, по отношению к которой голова на длинной и гибкой шее имеет значительную подвижность при ловле добычи и борьбе с нею.
Пищей завроптеригиям служили различные рыбы и головоногие (аммониты и белемниты). Большинство плезиозавров, обладавшие небольшой головой на тонкой и длинной шее, питались относительно некрупной добычей. Зато появившиеся в позднеюрское время крупные плиозавры (Pliosauroidea) имели огромный череп (до 3 м у 12-метровых ящеров) с острыми зубами до 10 см длиной. Эти морские хищники могли, вероятно, питаться крупными глубоководными головоногими подобно современным кашалотам и даже нападать на более мелких плезиозавров и других морских рептилий.
С завроптеригиями некоторые палеонтологи сближают также плакодонтов (Placodontia). Эта любопытная группа морских пресмыкающихся существовала в триасе. Некоторые плакодонты напоминали современных морских черепах и даже имели панцирь из остеодерм, другие - современных морских млекопитающих - сирен (дюгони и ламантины). Зубы плакодонтов имели форму широких массивных пластин и, вероятно, служили для раздавливания раковин моллюсков и панцирей ракообразных.
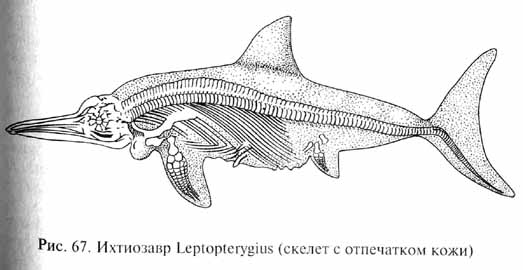
Наиболее совершенными пловцами среди морских рептилий мезозоя, бороздившими как прибрежные моря, так и открытый океан, были ихтиозавры (Ichthyopterygia). Их название означает "рыбоящеры", что подчеркивает внешнее сходство этих животных с рыбами. Тело ихтиозавров приобрело идеально приспособленную для быстрого плавания рыбообразную форму (рис. 67). Размеры ихтиозавров чаще всего составляли 3-5 м, но некоторые формы (например, Shonisaurus) достигали длины 18 м. Череп ихтиозавров напоминал череп дельфинов большими глазницами, длинной узкой мордой, у основания которой сверху располагались ноздри, и длинными тонкими челюстями, усаженными многочисленными тонкими и острыми зубами. Большинство ихтиозавров плавали за счет работы мощного хвостового плавника, расположенного, как у рыб, вертикально, спинной плавник служил стабилизатором движения. Однако скелет плавников у ихтиозавров был образован не костными или хрящевыми элементами, как у рыб, а плотной соединительной тканью, как у китообразных среди млекопитающих. Парные конечности, преобразованные в ласты, играли роль рулей глубины. У некоторых ихтиозавров, подобных Eurhinosaurus, парные конечности были сильно развиты и использовались как веслообразные движители при плавании. В ластах ихтиозавров, как и у плезиозавров, было значительно увеличено число фаланг пальцев, а у некоторых видов увеличено было и число пальцев (до 10), так что скелет конечности включал до 200 небольших костей, расположенных правильными рядами, обеспечивая прочность и гибкость ластов.
Развитие вторичного, т.е. не унаследованного от общих предков, сходства строения у разных видов организмов в результате формирования приспособлений к сходному образу жизни (т.е. сходному способу использования одних и тех же ресурсов внешней среды) получило название конвергенции. Одним из классических примеров конвергенции является развитие сходных признаков у ихтиозавров, дельфинов и рыб.
Ихтиозавры, вероятно, уже не могли выходить на сушу даже для откладки яиц. Возможно, эти рептилии были живородящими. Живорождение (даже с образованием примитивной плаценты) не так уж редко встречается среди пресмыкающихся (оно характерно для целого ряда видов современных ящериц и змей). О живорождении у ихтиозавров свидетельствуют некоторые палеонтологические данные: были обнаружены скелеты крупных особей (самок?), внутри которых находились скелетики маленьких ихтиозавров, иногда свернутые кольцом (как зародыши в яйце), иногда располагавшиеся так, что хвостовая часть скелета детеныша должна была выступать наружу из отверстия клоаки самки. Можно предполагать, что эти находки связаны с гибелью самки вместе с детенышем в момент родов. Судя по этим данным, у ихтиозавров при родах, как и у современных китообразных, детеныши выходили хвостом вперед, и сразу же рефлекторно начинал работать хвостовой плавник.
Любопытно, что вместе с некоторыми скелетами ихтиозавров были обнаружены остатки фоссилизованной кожи, причем сохранились даже пигментные гранулы. Возможно ихтиозавры имели темно-коричневую окраску. Было обнаружено также фоссилизованное содержимое желудков ихтиозавров, в нем оказались остатки головоногих моллюсков и рыб. Вероятно, в мезозойских морях ихтиозавры соответствовали в экологическом отношении современным дельфинам. Приспособление к сходному образу жизни в одной и той же среде обитания привело к значительному конвергентному сходству тех и других.
Происхождение специализированных водных рептилий - завроптеригий и ихтиозавров - до сих пор остается дискуссионным, поскольку, как и в большинстве случаев, ранние этапы эволюции этих групп не представлены в палеонтологической летописи. Некоторые палеонтологи склоняются к мнению об их независимом происхождении непосредственно от какой-либо группы котилозавров (капториноморфов или проколофонов). Высказывалось также предположение об общем происхождении завроптеригий и ихтиозавров от котилозавров или от примитивных синапсидных рептилий (пеликозавров), а в последние годы получила распространение гипотеза об их возникновении от примитивных диапсидных рептилий, близких к эозухиям (в этом случае принимают, что нижнее височное окно в их черепе вторично закрылось благодаря разрастанию костей).
Завроптеригии в лице своих наиболее примитивных представителей - нотозавров (Nothosauria), имевших еще не настоящие ласты, а лишь укороченные конечности с плавательными перепонками между пальцами, появились в раннем триасе. Ихтиозавры известны начиная с нижнего триаса. Расцвет обеих групп приходится на юрский период и первую половину мела.
Ископаемые остатки завроптеригий и ихтиозавров нередко встречаются в юрских осадочных породах на Русской равнине. Целые скелеты плиозавров были обнаружены при разработках горючих сланцев в Поволжье.
Помимо завроптеригий и ихтиозавров к жизни в море перешли и многие другие мезозойские рептилии. Можно сказать, что все подклассы пресмыкающихся имели в море своих представителей. В позднеюрское время появились первые морские черепахи. Архозавры были представлены в море специализированными группами крокодилов, конкурировавшими с ихтиозаврами. Морские крокодилы были очень многочисленны во второй половине мезозоя. Некоторые из них (метриоринхи - Metriorhynchidae), как и ихтиозавры, приобрели рыбообразное тело, вертикальный хвостовой плавник и ластообразные конечности.
Лепидозавры также делали неоднократные попытки "овладеть океаном". Среди морских ящериц наиболее замечательны позднемеловые мозазавры (Mosasauridae), достигавшие 10 м длины и ставшие столь же совершенными пловцами, как ихтиозавры и метриоринхи. Мозазавры по времени своего появления и существования были самыми поздними из этих трех конвергентных групп. Высказывались предположения, что конкуренция со стороны мозазавров могла сыграть роль в вымирании ихтиозавров, большинство видов которых исчезло уже в середине мела.
Век динозавров
К концу триаса произошло постепенное выравнивание климатических условий на значительной части суши, о чем говорит, в частности, очень однообразный и в целом сходный характер флоры на больших пространствах континентов, входивших прежде в состав Гондваны и Лавразии. Растительность юрского периода (рис. 68) характеризовалась преобладанием различных групп голосеменных (цикадовые, беннеттиты, хвойные, гинкговые) и древовидных папоротников; широко распространены были также крупные хвощи, среди которых, например, Equisetites arenaceus достигал 10 м высоты и 25 см в диаметре. Мезозойские хвойные были значительно разнообразнее и шире распространены, чем в наше время. В эту эру появились все современные семейства хвойных и большинство современных родов. Через Канаду, Сибирь и северную Часть Европы простирался пояс хвойных и листопадных лесов, произраставших в условиях умеренного климата. Южнее располагался еще более обширный пояс вечнозеленой субтропической растительности с обилием влажных и заболоченных местностей. Тропический экваториальный пояс был значительно шире современного.

В условиях ровного и очень теплого климата в средней части мезозоя завропсидные рептилии достигли максимального расцвета. Это было время удивительных животных, получивших широкую известность под названием "динозавры" ("страшные ящеры"). Название это относится к нескольким независимым группам рептилий из подкласса архозавров и в современном понимании не имеет определенного таксономического статуса (подобно термину "стегоцефалы", см. выше). Обычно динозавров распределяют в два отряда: ящеротазовых (Saurischia) и птицетазовых (Ornithischia), отличавшихся друг от друга рядом признаков. В частности, у них было разное строение тазового пояса: у птицетазовых лобковая кость имела особый отросток, тянувшийся назад под седалищной костью, тогда как у ящеротазовых такого отростка не было (рис. 69).

Динозавры одним или несколькими независимыми корнями (вопрос этот еще остается дискуссионным) произошли от псевдозухий. Древнейшие остатки ящеротазовых известны из средне-триасовых, а птицетазовых - из верхнетриасовых отложений Восточной и Южной Африки и Южной Америки. Примитивные динозавры унаследовали от псевдозухий тенденцию передвигаться на двух задних конечностях - бипедальность. В.Б.Суханов показал, что при сохранении примитивного положения конечностей (см. с.210) бипедальная локомоция позволяет увеличить скорость. Кроме того, при двуногом передвижении передняя часть тела высоко поднимается над почвой, что увеличивает обзор и уменьшает опасность перегрева при длительном пребывании животного на сухих открытых участках, где почва раскаляется под лучами яркого солнца. Бипедальность была широко распространена среди динозавров, хотя некоторые их группы по различным причинам вновь вернулись к опоре на четыре конечности.
Как и у высших звероподобных рептилий, у динозавров конечности располагались под телом в вертикальной плоскости, что обеспечивало большую экономичность передвижения по сравнению с группами, сохранившими позу пресмыкания. Вероятно, двуногие ящеры передвигались подобно современным страусоподобным птицам, используя бег или ходьбу с чередованием опоры на левую и правую ноги. Длинный мускулистый хвост уравновешивал высоко поднятую тяжелую голову и переднюю часть тела. Возможно, хвост служил также дополнительной опорой при стоянии в выпрямленной позе. Передние конечности у бипедальных динозавров либо использовались как хватательные, либо подвергались редукции.
В юрском и меловом периодах динозавры доминировали среди наземных позвоночных, заселив всевозможные местообитания и дав большое разнообразие форм, которые поражают воображение гротескной причудливостью облика, а иногда и огромными размерами.

Безусловный рекорд в последнем отношении принадлежит ящеротазовым динозаврам из группы завропод (Sauropoda), среди которых были самые крупные из когда-либо существовавших на Земле четвероногих животных: например, диплод
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





