|
ГЛАВА 4. КАЙНОЗОЙ - ВЕК МЛЕКОПИТАЮЩИХ И ПТИЦ
Физико-географические условия в начале палеогена мало отличались от таковых в конце мелового периода. После похолодания климат вновь стал теплым. В Европе до берегов Балтики восстановились тропические и субтропические флоры, имевшие характер густых и влажных лесов. В них были широко представлены пальмы, бананы, эвкалипты, вечнозеленые лавры, фикусы, магнолии, мирты, дубы, каштаны и другие широколиственные растения, а из хвойных - гигантские секвойи, араукарии, болотные кипарисы. В лесной тени произрастали папоротники, в том числе древовидные, по морским побережьям - тропическая мангровая растительность.
Тропические и субтропические флоры преобладали в начале палеогена в Северной и Южной Америках, Африке и Южной Азии. Даже в Гренландии и на Шпицбергене была разнообразная растительность, свойственная теплому умеренному климату: секвойи, болотные кипарисы, гинкго, буки, дубы, платаны, клены, магнолии, липы, березы и др. Любопытно развитие у многих высокоширотных растений гигантских листьев до 30-40 см. Климат в этих местах был в палеогене близок к современному климату Южной Франции или Калифорнии.
А на обширных территориях Центральной Азии с начала палеогена намечается тенденция к аридизации (повышениие засушливости) климата, которая, постепенно усиливаясь, прогрессировала на протяжении всего этого периода и неогена. Это привело к усыханию обильных здесь в позднемеловое время внутриконтинентальных пресноводных бассейнов, к постепенному обезлесиванию ландшафта с развитием к концу палеогена (олигоценовая эпоха) открытых пространств типа лесостепей, степей и саванн. Вместо влаголюбивой сочной лесной растительности получают преобладание злаки (Gramineae), жесткие и относительно сухие листья которых хорошо защищены от обезвоживания. Местами в саваннах возвышались отдельные группы деревьев с высоко расположенными кронами. Более густая древесная и кустарниковая растительность сохранялась по долинам рек. Аналогичные открытые сухие ландшафты получили в начале неогена широкое распространение и на других континентах, но наиболее засушливыми в кайнозое оставались районы Центральной Азии.
Во второй половине палеогена началось постепенное глобальное похолодание климата, которое усилилось в неогене. Оно завершилось в плейстоценовой эпохе четвертичного периода развитием нового великого оледенения. Процесс похолодания климата был очень медленным и постепенным. Все более отчетливо проявлялась климатическая зональность. Климатические изменения легко прослеживаются по преобразованиям флоры. Теплолюбивая флора в Евразии и Северной Америке постепенно отступала к югу, сменяясь в умеренной зоне листопадными и хвойными лесами. Изменения климата представляют собой внешний итог сложнейшего взаимодействия множества различных факторов (космических, географических и атмосферных), среди которых важную роль играют направления морских течений и характер атмосферной циркуляции. Эти последние зависят, в свою очередь, от изменений рельефа и положения континентов.
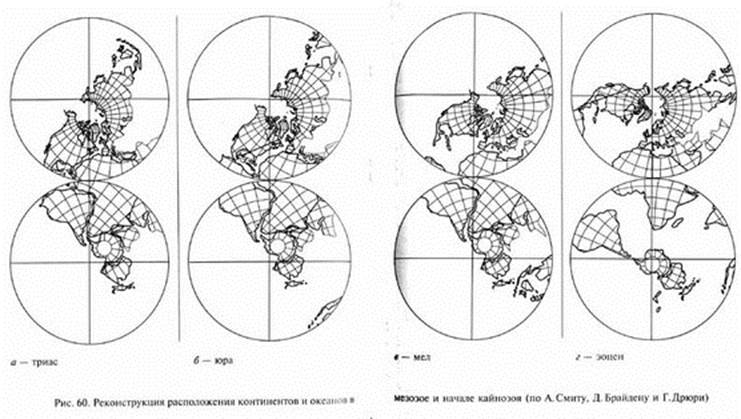
Согласно представлениям теории тектоники плит, в кайнозое продолжалось движение континентов, которое постепенно привело к современной картине их размещения. От Северо-Американского материка отделилась Гренландия. Смещения Африки и Индии почти совсем закрыли мезозойский океан Тетис. На его месте ныне существуют Средиземное, Черное и Каспийское моря, а на востоке - моря Зондского архипелага, однако среди ученых еще нет единства мнений о преемственности этих бассейнов по отношению к Тетису. Индостанская плита, смещаясь к северо-востоку, наконец, соединилась с Азией. Австралия, вероятно, лишь в эоцене обособилась от Антарктиды и стала быстро (в геологическом смысле) продвигаться к северу, приближаясь к экватору (см. рис. 60).
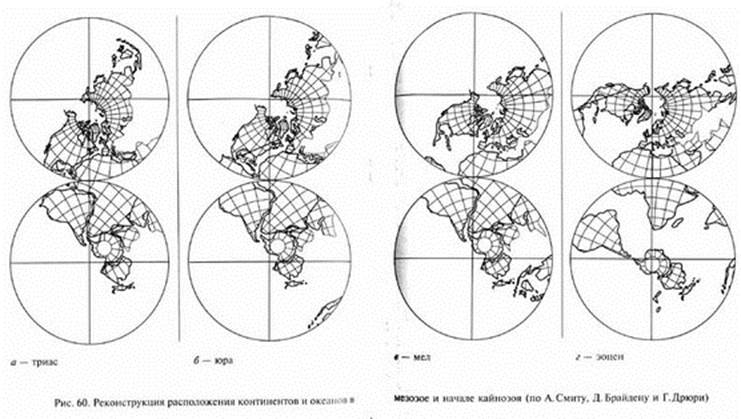
Смыкание континентальных плит сопровождалось развитием горной складчатости в зонах их контакта. В кайнозое сформировался Альпийский складчатый пояс. В палеогене и неогене поднялись горные цепи Атласа, Пиренеев, Альп, Апеннин, Балкан, Карпат, Крыма, Кавказа, Копетдага, Памира и Гималаев, завершилось формирование Анд, Кордильер и ряда других горных систем. Неоген характеризуется высоким положением материков И наибольшей степенью осушения их территорий за весь фанерозой, с этим связано преобладание открытых сухих ландшафтов в неогене.
Северный полюс, находившийся в мезозое в северной части Тихого океана и на близких к океанскому побережью областях Северо-Восточной Азии и Аляски, в кайнозое оказался в центре почти замкнутого бассейна Северного Ледовитого океана, который сам сформировался лишь в кайнозое. Этот бассейн в значительной своей части закрыт для доступа меридиональных морских течений, которые могли бы обеспечить более равномерное распределение температур в мировом океане и на поверхности Земли в целом. Благодаря этому Северный Ледовитый океан при расположении в нем Северного полюса оказался аналогичным континенту в смысле возникновения там огромного "очага холода", "мешка льдов". Возможно, это было важнейшей из причин, вызвавших охлаждение климата в кайнозое, которое завершилось великим оледенением.
Первый обширный ледниковый щит сформировался в Антарктиде в миоценовую эпоху неогена около 20 млн лет назад. До этого в течение палеогена в Антарктиде сохранялся умеренный прохладный климат, и ледники имелись лишь в горах. Развитие ледникового щита Антарктиды вместе с постепенно расширявшейся зоной плавучих льдов в Северном Ледовитом океане значительно увеличили отражение солнечных лучей (альбедо) земной поверхности, что, в свою очередь, стало фактором, способствовавшим дальнейшему охлаждению климата. Климатическое равновесие все более нарушалось в сторону увеличения контрастов климатических зон. Наконец, в плейстоценовой эпохе четвертичного периода (около 3 млн, а по некоторым данным, даже 1-5 млн лет назад) началось великое оледенение, сопровождавшееся развитием гигантских ледниковых щитов в Европе и Северной Америке. Оледенение прерывалось несколько раз резко наступавшими потеплениями (межледниковые эпохи, или интерстадиалы), во время которых ледниковый покров на континентах Европы и Северной Америки стаивал практически полностью. Затем происходило новое понижение температур с последующим оледенением. Период этих изменений составлял в среднем около 100 тыс. лет. Такой характер процесса изменений климата говорит за то, что развитие оледенения в четвертичном периоде носит пороговый характер: при незначительном нарушении неустойчивого температурного баланса, когда средние температуры падают ниже некоторого порогового уровня, начинает быстро прогрессировать оледенение и наоборот. Среди причин, влиявших на колебания климата в ту или другую сторону, указывали на вулканизм (снижение прозрачности атмосферы в результате ее засорения вулканическим пеплом), а также на колебания наклона экватора Земли к плоскости ее орбиты и изменения параметров самой орбиты под действием притяжения других планет.
Максимальное из плейстоценовых оледенений (около 250 тыс. лет назад) покрывало территорию около 45 млн км (т. е. примерно 30% суши), ледниковый шит в Европе достигал широты 48° 30', а в Северной Америке - 37°. Объем льдов на суше в это время составлял около 55 млн км3. Соответственно уровень мирового океана в эпохи оледенений значительно падал (на 85-120 м), и возникали сухопутные связи между континентами и островами, разобщенными ныне проливами. Например, Берингийская суша связывала в эпохи оледенений Аляску и Чукотку, Британские острова соединялись с Европой и т.д. Последнее оледенение завершилось около 10 тыс. лет назад.
Развитие жизни в палеогене
В начале палеоценовой эпохи фауна млекопитающих оставалась, по существу, той же, что и в позднемеловое время. В ней были представлены лишь группы, возникшие еще в мезозое: растительноядные многобугорчатые, внешне напоминавшие грызунов, но, возможно, родственные прототериям - однопроходным, а также архаические представители сумчатых и плацентарных, питавшиеся насекомыми и другой мелкой добычей. Для всех архаических млекопитающих были характерны такие примитивные особенности, как относительно небольшой мозг, простые треугольные зубы (за исключением многобугорчатых), пятипалые конечности, опиравшиеся при передвижении на всю кисть и стопу (плантиградность).
Предки млекопитающих - синапсидные рептилии-териодонты вымерли еще в позднем триасе. Однако в 1992 г. было опубликовано сенсационное сообщение о находке в позднепалеоценовых отложениях Альберты в Канаде фрагмента нижней челюсти, ряд особенностей которой очень сходен с состоянием цинодонтов. Если данные об этом палеоценовом цинодонте, получившем название Chronoperates, подтвердятся, придется заключить, что одна из филетических линий териодонтов просуществовала весь мезозой и перешла в кайнозой, пережив (вместе с некоторыми линиями динозавров) рубеж мела и палеогена. (Однако хроноператес может оказаться не цинодонтом, а представителем одной из очень архаических мезозойских групп млекопитающих - симметродонтов, сохранившим дополнительные кости в нижней челюсти).
К середине палеоценовой эпохи разнообразие млекопитающих значительно возросло и настолько, что можно предположить расхождение их предковых линий еще до конца мела. Но основная адаптивная радиация плацентарных и сумчатых происходила в палеоцене и эоцене, когда сложились все основные отряды кайнозойских млекопитающих. (При обсуждении филогении различных отрядов плацентарных в этой главе не будут рассматриваться приматы, эволюционная история которых разбирается в следующей главе).
От примитивных насекомоядных плацентарных возникли всеядные (использующие как животную, так и растительную пищу), а затем и настоящие растительноядные формы. Растительноядность у некоторых групп плацентарных развилась в палеоцене. Начало этого направления приспособительной эволюции было представлено архаическими копытными - кондиляртрами (Condylarthra), древнейшие представители которых известны из верхнемеловых отложений Южной Америки. Это были сравнительно небольшие животные размером с зайца, более поздние представители достигали длины тела около 180 см. Внешне они больше походили на хищников, чем на копытных, и сохраняли большие клыки. Однако их коренные зубы имели хотя и низкие, но широкие коронки, пригодные для перетирания не особенно жесткой растительной пищи. Ключицы были утрачены, что говорит о приспособлении локомоторного аппарата к быстрым "монотонным" движениям конечностей при беге, а концевые фаланги пятипалых кисти и стопы были защищены небольшими копытцами (по крайней мере у некоторых форм, подобных Phenacodus). Вероятно, при беге конечности этих животных опирались уже не на всю стопу, а лишь на пальцы. Возможно, примитивные кондиляртры были предками других групп копытных.
Среди этих последних уже в позднем палеоцене и эоцене появились более специализированные, крупные и причудливые формы. Характерны в этом отношении диноцераты (Dinocerata - "страшнорогие"), которые были самыми крупными наземными млекопитающими эоценовой эпохи, достигая размеров современных носорогов. Относительно короткие и толстые пятипалые конечности этих массивных животных несли копыта. Череп у некоторых форм (например, уинтатерия - Uintatherium, рис.80) имел рогоподобные костные выросты и кинжалообразные острые клыки. Вероятно, эти крупные животные были хорошо защищены от нападений современных им хищников. Однако диноцераты вымерли уже к концу эоцена. Скорее всего, их вымирание связано с конкуренцией с более прогрессивными группами копытных, которую диноцераты проиграли благодаря общему консерватизму своей организации, в частности сохранению относительно очень небольшого головного мозга.

В палеоцене и эоцене появились такие прогрессивные группы растительноядных млекопитающих, как непарнокопытные (Perissodactyla), парнокопытные (Artiodactyla), грызуны (Rodentia), зайцеобразные (Lagomorpha) и ряд других. Их быстрая адаптивная радиация привела к вымиранию архаических групп растительноядных зверей: кондиляртр, диноцерат и родственных последним пантодонтов (Pantodonta) к концу эоцена, многобугорчатых в раннем олигоцене.
Своеобразным убежищем для некоторых примитивных групп млекопитающих стала Южная Америка, которая к концу раннего эоцена обособилась от Северной Америки и оставалась изолированной до плиоцена. В палеогене и неогене там существовал удивительный мир причудливых форм, сохранявших общий примитивный уровень организации; их судьбу мы рассмотрим несколько ниже. Среди высших копытных раньше других начали адаптивную радиацию непарнокопытные, которые уже в эоцене были представлены разнообразными формами. Центром эволюции этого отряда была, по-видимому, Северная Америка, откуда известны наиболее ранние и примитивные представители различных семейств, как доживших до современности (лошади, тапиры, носороги), так и вымерших (титанотерии, халикотерии и др.). Непарнокопытные пережили свой расцвет в палеогене, причем их эволюционная история стала одной из самых ярких страниц в кайнозойской палеонтологической летописи.
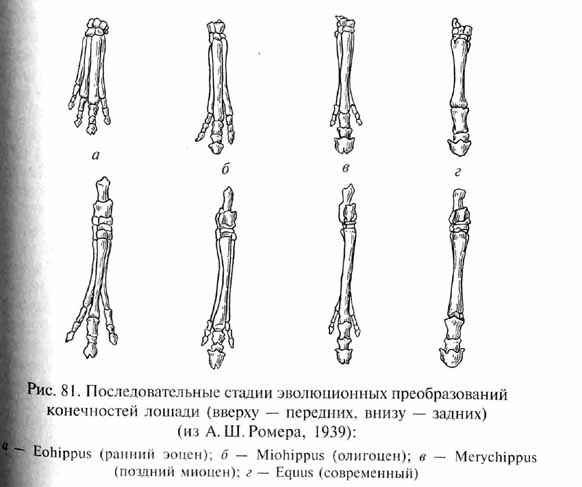
Примитивные эоценовые непарнокопытные были небольшими животными, у которых уже наметились изменения конечностей, связанные с их приспособлением к быстрому бегу, в частности, удлинение средних пальцев и элементов пястья и плюсны при одновременной редукции боковых лучей в кисти и стопе. У эоценовых форм было по 4 пальца на передних и по 3 на задних конечностях (такое состояние сохранилось у современных тапиров). Эволюционная тенденция к развитию тонких удлиненных конечностей как приспособление к убыстрению бега наиболее ярко выражена в филогенезе лошадей (Equidae, рис. 81), который является одним из наиболее изученных. Близко к основанию филогенетического ствола лошадиных стоит раннеэоценовый эогиппус (Eohipipus). Это животное размерами и пропорциями тела напоминало небольшую собаку (длина тела составляла около 50 см). Вероятно, эогиппусы жили в лесных зарослях и питались сочной и мягкой растительностью. В филогенезе лошадей происходило постепенное увеличение размеров тела, сопровождавшееся коррелятивным удлинением морды. Эти тенденции проявились уже у позднеэоце-новых (Orohippus, Epihippus) и олигоценовых (Ме8огирри5, Miohip-tous) лошадей. Последние достигали размеров более 1 м и сохранили лишь по 3 пальца на всех конечностях. Они оставались лесными животными. Дальнейшая эволюция лошадей связана с приспособлением к жизни в открытых местообитаниях, широко распространившихся в неогене, и будет рассмотрена нами ниже. Тенденция к увеличению размеров тела еще сильнее была выражена у титанотериев и носорогов, эволюция которых также начиналась с небольших легко построенных эоценовых форм. Но уже в олигоцене эти непарнокопытные были представлены огромными животными, с размерами которых могут сравниться среди наземных млекопитающих лишь неогеновые хоботные.
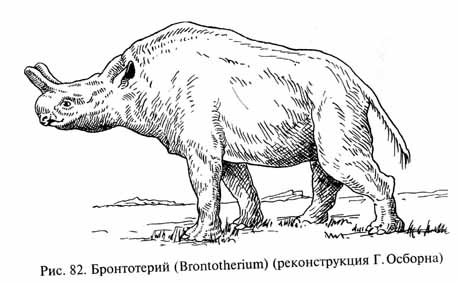
Титанотерии или бронтотерии (Bronthotheriidae, рис. 82) достигали в холке высоты около 2,5 м при длине тела до 4,5 м. Их длинный и низкий череп с поразительно маленькой мозговой полостью имел на носовых или на лобных костях большие рогоподобные выступы - парные или в виде толстого срединного "рога", раздвоенного у вершины. Титанотерии обитали в лесах, на влажных лугах вблизи водоемов, а некоторые виды вели, вероятно, полуводный образ жизни. Питались они сочной лесной или водной растительностью. Титанотерии вымерли уже в раннем олигоцене. Их заместили представители другой группы непарнокопытных - носороги (Rhinocerotoidea), пять видов которых дожили до нашего времени. В олигоцене же и в неогене это была процветающая и очень разнообразная группа.
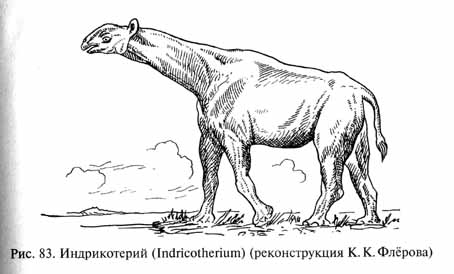
В отличие от современных носорогов, несущих на морде один или два рога (представляющих собой, как это ни кажется странным, пучки сросшихся волос), многие древние носороги были безрогими (название этой группе было дано по современным формам). Среди них имелись относительно легкие, так называемые "бегающие носороги" - гиракодонты (Hyracodontidae), полуводные, внешне несколько напоминавшие бегемотов аминодонты (Amynodontidae), но особенно замечательны были гигантские носороги (Indricotheriidae), распространенные в Азии в олигоцене в начале миоцена. Это были крупнейшие наземные млекопитающие, превосходившие даже крупных хоботных и уступавшие размерам и массе тела лишь завроподам: длина их тела достигала 7-8 м при высоте в холке до 5,5 м. Довольно длинная и мощная шея могла поднять огромную голову (череп имел до 1,2 м в длину) на высоту около 8 м над землей (рис. 83). Гигантские носороги обитали в открытых ландшафтах, лесостепях и саваннах, которые были широко распространены в Азии уже в олигоцене. Вероятно, эти животные питались листвой деревьев с высокими кронами. Дальнейшая аридизация азиатских степей в неогене привела к их вымиранию.
Заслуживает упоминания еще одна интересная группа непарнокопытных, просуществовавшая от позднего эоцена до раннего плейстоцена - халикотерии (Chalicotheriidae). По размерам тела и форме головы эти животные несколько напоминали лошадей, но их трех-четырехпалые конечности были снабжены не копытами, а большими изогнутыми когтями, похожими на когти гигантских ленивцев (см. ниже) и муравьедов. Любопытно, что создатель палеонтологии Ж.Кювье в свое время описал когтевую фалангу халикотерия как принадлежащую муравьеду, будучи уверен в существовании жестких и однозначных корреляций между различными частями организма. Среди палеонтологов нет единства мнений По поводу образа жизни этих странных существ. Возможно, халикотерии, подобно гигантским ленивцам, питались молодыми побегами и листьями деревьев, ветви которых они подгибали и удерживали когтистыми лапами. По другому предположению, халикотерии использовали когти для выкапывания из почвы питательных Клубней и корневищ растений.
Адаптивная радиация парнокопытных началась несколько по-3ДНее, чем непарнокопытных, однако в олигоцене уже существовали примитивные представители современных семейств свиных (Suidae), верблюдов (Camelidae), оленьков (Tragulidae) и ряд вымерших групп. Анализируя филогенетические преобразования скелета конечностей у палеогеновых парнокопытных, В.О.Ковалевский разработал концепцию об адаптивных и инадаптивных изменениях органов в эволюции (см. ч. II, гл. 5).
Эволюция парно- и непарнокопытных в палеогене происходила на территориях Северной Америки и Евразии, связанных друг с другом Берингийским сухопутным "мостом".
В Африке сформировалась в палеогене своя оригинальная фауна растительноядных млекопитающих. В ее состав входили разнообразные животные: крупные хоботные (Proboscidea); небольшие даманы, или жиряки (Hyracoidea), напоминавшие своим внешним обликом и образом жизни некоторых грызунов; вымершие еще в палеогене эмбритоподы (Embrithopoda), внешне сходные с носорогами и титанотериями; наконец, перешедшие к полуводному и водному образу жизни десмостилы (Desmostylia), несколько напоминавшие давно вымерших плакодонтов, и сиреновые (Sirenia), у которых развился хвостовой плавник, передние конечности преобразовались в ласты, а задние редуцировались. При всем внешнем несходстве этих групп животных друг с другом ряд характерных общих признаков (увеличение одной пары резцов, строение коренных зубов, редукция ключиц, наличие копыт, положение млечных желез и др.) позволяют предполагать их общее происхождение. Некоторые авторы считают, что их предками могли быть примитивные кондиляртры (однако в Африке, где возникли все эти группы, ископаемых остатков кондиляртр пока не обнаружено).
Древнейшим представителем хоботных является среднеэоце-новый нумидотерий (Numidotherium). Более известен меритерий (Moeritherium) из верхнего эоцена - нижнего олигоцена, который размерами и внешним обликом, вероятно, напоминал тапира и имел зачаточный хобот. Вторые пары резцов в верхней и нижней челюстях были сильно увеличены; их дальнейшая гипертрофия у более поздних хоботных привела к формированию бивней.
В раннем олигоцене появились древнейшие мастодонты - более высокоразвитые хоботные, внешне уже напоминавшие настоящих слонов. Название "мастодонт" означает буквально "сосце-зуб", что связано с характерной формой увеличенных коренных зубов, коронки которых имели сосцевидные бугорки. По паре гипертрофированных резцов-бивней было как в верхней, так и в нижней челюсти. Морда у мастодонтов оставалась удлиненной, поэтому хобот, представляющий собой результат разрастания верхней губы и носа, был относительно коротким. Мастодонты были много крупнее меритерия, некоторые виды уже в олигоцене достигали размеров современных слонов. Расцвет хоботных наступил уже в неогене.
Сирены, в современной фауне представленные лишь двумя родами (ламантином - Trichechus и дюгонем - Dugong), обособились, возможно, еще в палеоцене в связи с приспособлением к постоянной жизни в воде. Сирены возникли в прибрежных водах теплых морей еще существовавшего тогда океана Тетис и были широко распространены уже в эоцене. В отличие от сирен десмостилы, обитавшие в олигоцене и миоцене по берегам Тихого океана, сохранили обе пары конечностей и могли передвигаться по суше, хотя и очень неуклюже, так как их конечности были широко расставлены в стороны. Характерно строение их массивных коренных зубов, образованных 4-8 дентиновыми трубками, покрытыми тонким слоем эмали, - возможно, эти зубы служили для раздавливания раковин моллюсков.
В палеогеновых морях помимо сирен появились и другие млекопитающие, которые заняли освободившиеся экологические ниши морских рептилий. Это были древнейшие китообразные (Cetacea), остатки которых обнаружены в среднеэоценовых отложениях Северной Африки, т. е. также в области эпиконтиненталь-ных морей океана Тетис. Пропорции тела архаических китов (Archaeoceti) значительно отличались от таковых у современных групп: туловище было длинное и относительно тонкое (у наиболее крупной формы, зейглодона - Zeuglodon - до 21 м в длину при диаметре не более 2,5 м). Длинные узкие челюсти большого, до 1,2 м в длину, и низкого черепа были усажены впереди коническими, а сзади сжатыми с боков острыми зубами. Короткие передние конечности располагались близко к голове, а задние были сильно редуцированы (полностью развитые задние конечности сохранялись у эоценовых Ambulocetus и Basilosaurus). По всей вероятности, китообразные возникли от каких-то примитивных наземных хищников, перешедших к рыбоядности и постепенно развивавших приспособления к амфибиотической, а потом и к водной жизни. Примитивные представители современных групп китообразных: зубатых (Odontoceti) и беззубых (Mystacoceti) китов появились соответственно в позднем эоцене и олигоцене. (По мнению некоторых палеонтологов, происхождение китообразных связано не с хищными, а с древнейшими парнокопытными).
Морская ихтиофауна палеогена уже включала все современные крупные группы рыб, хотя видовой состав и отличался от современного. Уже в плиоценовой эпохе неогена существовала крупнейшая из хищных акул Megalodon, длина тела которой достигала 30 м, а диаметр открытой пасти был свыше 2 м.
Вернемся, однако, на сушу, где разворачивались наиболее интересные события кайнозойской эволюции органического мира. До сих пор мы концентрировали внимание на растительноядных млекопитающих, в связи с их важной ролью в биоценозах как первичных потребителей растительной биомассы. От примитивных насекомоядных предков возникли также разнообразные хищники, широко представленные среди палеогеновых млекопитающих.
Древнейшие представители отряда плацентарных хищников (Carnivora) - креодонты (Creodonta) по некоторым чертам своей организации были близки к примитивным копытным - кондиляртрам и, вероятно, имели общее с ними происхождение. Креодонты разделяли все примитивные особенности архаических групп млекопитающих (длинный низкий череп с маленькой мозговой коробкой, пятипалая конечность, плантиградность и др.)- Ископаемые остатки креодонтов известны, начиная с нижних горизонтов палеоцена, поэтому можно полагать, что обособление их эволюционного ствола произошло еще до конца мела. Наиболее ранние креодонты имели небольшие размеры, тонкие конечности и стройное тело, представляя переходный тип от насекомоядных предков. Однако уже в палеоцене появились более крупные хищники, среди которых арктоцион (Arctocyon) размерами и телосложением напоминал медведей. В раннем олигоцене существовали еще более крупные и тяжело построенные креодонты; самый большой из них, эндрюсархус (Andrewsarchus), достигал длины около 4 м при высоте в холке до 2 м, его череп имел длину 0,9 м. И крупные, и мелкие креодонты питались различными архаическими растительноядными млекопитающими и другой доступной им добычей. Поведение креодонтов было, вероятно, относительно простым по сравнению с поведением современных хищников как в отношении приемов охоты, так и в уходе за потомством.
После возникновения новых групп млекопитающих с более высоко развитым головным мозгом креодонты должны были сойти со сцены. Уже из верхнеэоценовых отложений известны ископаемые остатки древнейших представителей высших хищников (Fissipedia), обладавших сравнительно большим головным мозгом и рядом других прогрессивных признаков. В олигоцене последние креодонты вымирают.
В палеогене помимо млекопитающих процветали и многие другие группы наземных позвоночных. Во влажных тропических лесах, в пресных водоемах в изобилии встречались бесхвостые И хвостатые земноводные; в более открытых местообитаниях были многочисленны ящерицы и змеи. Число чешуйчатых рептилии в кайнозое ничуть не уменьшилось, и возникли некоторые новые семейства (например, в палеогене сформировались высшие змеи - Caenophidia, к которым принадлежат полозовые, или ужовые, - Colubridae, и все группы ядовитых змей). На суше, в пресных водоемах и в морях жили разнообразные черепахи. Представители единственной группы архозавров, пережившей великое вымирание в конце мела - крокодилы - процветали в жарком климате палеогена и были широко распространены на всех континентах. Наконец, потомки архозавров, птицы в кайнозое стали самой обильной видами группой наземных позвоночных.
Между птицами и млекопитающими произошел своего рода "раздел сфер влияния": большинство видов зверей, возникших от сумеречных и ночных мезозойских млекопитающих, сохранили и в кайнозое предпочтение к ночной активности, тогда как птицы, потомки гелиотермных рептилий, остались преимущественно дневными формами. Например, в составе современной фауны тропических лесов Габона и Панамы дневной образ жизни ведет 96 % видов птиц, тогда как для 70% видов млекопитающих характерна ночная активность. Разумеется, оба класса высших амниот отчасти проникли и в "чужую сферу влияния": имеются преимущественно дневные группы млекопитающих и ночные птицы. Но это относится к значительному меньшинству видов в обоих классах и потребовало развития у них специальных адаптации, не характерных для данного класса в целом. У млекопитающих, по мнению Ш.-Д.Пьера, с освоением дневной активности связано увеличение размеров тела и развитие специализированной пищеварительной системы, позволяющей использовать относительно грубые, трудно перевариваемые и объемные растительные корма (напомним о крупных и очень крупных животных среди растительноядных млекопитающих). Интересно появление в раннем эоцене крупных нелетающих наземных птиц с редуцированными крыльями. Некоторые из них обладали высоким массивным клювом с берыми краями и были, вероятно, опасными хищниками. Эоценовые диатримы (Diatryma. рис. 84), Достигавшие двухметровой высоты, или более поздние (олигоцен-плейстоцен) и еще более крупные (до 3 м) фороракосы (Phororhacidae) напоминали двуногих хищных динозавров (разумеется, на более высоком уровне организации птиц). Существование таких крупных нелетающих птиц показывает, что после вымирания динозавров птицы, как и млекопитающие, сделали попытку занять освободившиеся в биоценозах экологические ниши крупных хищников. Неудача птиц в этой роли может быть связана с меньшим по сравнению со зверями совершенством размножения и трудностями насиживания яиц и выращивания птенцов для нелетающих видов, численность которых как крупных хищников не могла быть высокой. Во всяком случае, эти своеобразные "пернатые динозавры" вымерли после появления высших групп хищных млекопитающих.

Вероятно, еще в меловое время от остальных птиц обособились нелетающие страусоподобные птицы - группа бескилевых (Ratitae), ископаемые остатки которых известны из палеоцена Бразилии (Diogenornis) и среднего эоцена Германии (Palaeotis). Возможно, бескилевые птицы представляют собой ряд независимых филогенетических стволов, эволюционировавших параллельно друг другу. Утрата способности к полету происходила на разных этапах эволюционной истории птиц у многих групп и была, вероятно, связана с отсутствием опасных хищников в местообитаниях, занимаемых птицами. Не случайно нелетающие птицы часто возникали на островах и изолированных континентах (Австралия, Южная Америка).
Уже с позднеэоценового времени известны пингвины (Impen-nes), ископаемые остатки которых обнаружены в Новой Зеландии и Южной Америке. Вся эволюционная история этой группы протекала в Южном полушарии. Высказывались предположения, что уход пингвинов с суши в океан связан с оледенением Антарктиды, но это неверно: пингвины как группа морских рыбоядных птиц, утративших способность к полету, сформировались задолго до начала оледенения (и среди современных пингвинов многие виды гнездятся в умеренной зоне, а один вид - даже в тропиках). Ближайшими родственниками пингвинов являются хорошо летающие трубконосые птицы (Procellariae), к которым относятся альбатросы и буревестники.
На отрезанных материках
Мы уже упоминали, что Австралия, Южная Америка и Антарктида по крайней мере с эоцена полностью обособились друг от друга и от всех других континентов широкими морскими проливами и морями. К. сожалению, о развитии жизни в Антарктиде перед началом последнего оледенения известно очень мало.
Австралия уже со времени распада Гондваны занимала обособленное положение, будучи связана с другими материками, главным образом, через Антарктиду (см. рис. 60). Поэтому в Австралию ко времени ее изоляции, вероятно, еще не проникли плацентарные млекопитающие. В Австралии сформировалась уникальная фауна сумчатых и однопроходных, отчасти сохранившаяся до наших дней. Однако палентологическая история австралийских млекопитающих известна еще очень плохо. Вероятно, современная фауна этого материка значительно обеднена по сравнению с предшествовавшими эпохами кайнозоя. Относительно хорошо известна поздняя - плейстоценовая - фауна Австралии. Среди плейстоценовых сумчатых здесь можно упомянуть дипротодона (Diprotodon) - самого крупного среди известных сумчатых, достигавшего размеров носорога с черепом около 1 м и мощными бивнеподобными резцами в верхней и нижней челюстях. Очень своеобразен был "сумчатый лев" (Thylacoleo), у которого коренные зубы были редуцированы, зато имелись огромные режущие предкоренные. Некоторые палеонтологи считают
;это животное хищником, другие же полагают, что "сумчатый лев" питался какими-то плодами.
В Южную Америку до ее отделения от Северной Америки успели проникнуть некоторые архаические группы плацентарных млекопитающих, имелись там и сумчатые. На основе этих групп в течение длительной изоляции Южной Америки сложилась удивительная фауна архаических млекопитающих. В Северной Америке, Европе и Азии в течение палеогена и неогена плацентарные млекопитающие в целом сделали большой шаг вперед по пути морфофизиологического прогресса (увеличение и усовершенствование головного мозга, усложнение поведения, прогрессивные изменения локомоторного аппарата, органов пищеварения, зубной системы и т. д.). В Южной Америке все это время продолжали процветать архаические группы, сохранявшие примитивное строение головного мозга, но также на свой лад совершенствовавшие органы пищеварения и локомоции, причем многие их адаптации оказались очень сходными с соответствующими приспособления-Ми млекопитающих Старого Света и Северной Америки.
Очень разнообразны и причудливы были южноамериканские растительноядные млекопитающие. Копытные были представлены группами нотоунгулят (Notoungulata), астрапотериев (Astra-Potheria), литоптерн (Litopterna) и пиротериев (Pyrotheria). Нотоунгуляты известны также из палеоцена Азии (где, вероятно, и возникла эта группа) и раннего эоцена Северной Америки, но исчезли повсеместно уже в эоцене, сохранившись и процветая лишь в Южной Америке. Южноамериканские нотоунгуляты были очень разнообразны; среди них были и проворные мелкие животные, подобные типотериям (Typotheria), с длинными задними конечностями, которые напоминали кроликов пропорциями тела, размерами и строением зубов; и тяжелые коротконогие токсо-донты (Toxodonta), достигавшие 2,75 м длины и похожие на некоторых безрогих носорогов.
Астрапотерии и пиротерии были крупнейшими копытными Южной Америки, достигая размеров крупных носорогов и даже слонов. С последними они были сходны также строением конечностей и наличием хобота, а пиротерии удивительно напоминали настоящих хоботных еще и обликом черепа и зубной системы (вплоть до крупных бивнеподобных резцов в верхней и нижней челюстях).
В эволюции литоптерн возникли формы, конечности которых были поразительно сходны с лошадиными (яркий пример эволюционного параллелизма). В процессе приспособления к быстрому бегу по относительно плотной почве саванн у этих литоптерн усиливался средний палец и редуцировались боковые (рис. 85), вплоть до полной их редукции у миоценового тоатерия (Thoatherium). Тоатерий был в этом отношении даже более специализирован, чем настоящие лошади, сохраняющие рудименты боковых пальцев в виде так называемых грифельных косточек (см. рис. 81). Наиболее поздний представитель литоптерн - плейстоценовая макраухения (Macrauchenia) размерами и пропорциями тела напоминала верблюда, но имела трехпалые ноги, а ноздри на черепе располагались сверху, как у хоботных животных (возможно, макраухения также имела небольшой хобот).

Таким образом, при отсутствии в Южной Америке настоящих хоботных, носорогов, лошадей, зайцеобразных и других групп млекопитающих, эволюционировавших в Евразии и Северной Америке, там конвергентно возникли формы, удивительно напоминавшие внешне своих аналогов.
Зато совершенно специфичны южноамериканские неполнозубые (Edentata). Эта группа, возможно, возникла в Северной Америке, откуда известны остатки древнейших палеоценовых неполнозубых, но центром ее эволюции стала Южная Америка в условиях географической изоляции. И современные неполнозубые (ленивцы, броненосцы, муравьеды) обитают в основном в Южной Америке, за исключением нескольких видов, проникших в южную часть Северной Америки после восстановления ее связи с Южной. Среди вымерших неполнозубых интересны родственные броненосцам глиптодонты (Glyptodontidae, рис. 86), достигавшие до 5 м в длину. Их тело было одето сплошным панцирем из сросшихся мелких костных пластинок - остеодерм. "Шапочка" из остеодерм прикрывала сверху голову. Пальцы коротких мощных лап были защищены копытами.


Но особенно интересной группой неполнозубых были гигантские наземные ленивцы, или мегатерии (Megatheriidae, рис. 87). Эти животные, как и глиптодонты, известны начиная с олигоце-на. Некоторые более поздние представители наземных ленивцев достигали огромных размеров: плейстоценовый мегатерий (Megatherium) был высотой со слона при длине тела около 6 м. Внешний облик этих странных существ был необычен. Огромное тело опиралось на мощные конечности, особенно велики были задние лапы с большими широкими ступнями. Передние конечности несли крупные изогнутые когти и при ходьбе опирались не на ладонную, а на боковой край тыльной стороны, сгибая пальцы внутрь. Веро
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





