|
ГЛАВА 1. ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ И ЭВОЛЮЦИЯ
Между мутациями и их проявлением в фенотипе лежат сложные процессы морфогенеза, в ходе которых наследственная информация, преобразуясь, переходит на все более высокие уровни (молекулы - клетки - ткани - органы). В то же время развивающийся организм подвергается воздействию различных внешних Факторов, направляющих процессы морфогенеза в те или другие русла, приводя к формированию конкретных фенотипических особенностей. Сущность этих процессов и механизмы преобразования информации известны еще сравнительно слабо; обобщающая теория индивидуального развития еще не создана. Это очень затрудняет использование эмбриологических концепций в эволюционистике. Данные и обобщения онтогенетики еще не заняли подобающего места в синтетической теории эволюции.
Дифференцировка организма в онтогенезе
Одной из основных проблем онтогенетики, решение которой должно сыграть важную роль и для эволюционной теории, остается сущность явлений онтогенетической дифференцировки, т. е. тех процессов, которые ведут к развитию разнокачественности различных фенотипических структур, возникающих в онтогенезе, в конечном итоге, из единственной клетки - зиготы. Даже в организации столь просто устроенных животных, как кишечнополостные, насчитывается около 10 разных типов клеток, а в организме млекопитающих - порядка 1000 типов клеток. Клетки разных типов группируются в ткани и формируют многотканевые комплексы со сложной пространственной структурой - органы.
По современным представлениям, в процессе дробления зиготы на бластомеры и при последующих делениях соматических клеток каждая дочерняя клетка в норме получает 100 % наследственной информации, т.е. полный набор генов, точно соответствующий исходному генотипу зиготы. Однако соматические клетки резко различаются по строению и функциям (достаточно сравнить, например, нейрон, мышечную клетку и эритроцит в одном организме).
Можно считать установленным, что различия соматических клеток определяются дифференциальной активностью генов (другими словами, в одних типах клеток активны одни, а в других - другие участки хромосом). Неактивные в данной клетке гены блокируются какими-то входящими в состав хромосом веществами, какими именно - еще точно не выяснено. Предполагают, что этими веществами (репрессорами) являются те или иные хромосомные белки (многие ученые считают репрессорами белки-гистоны).
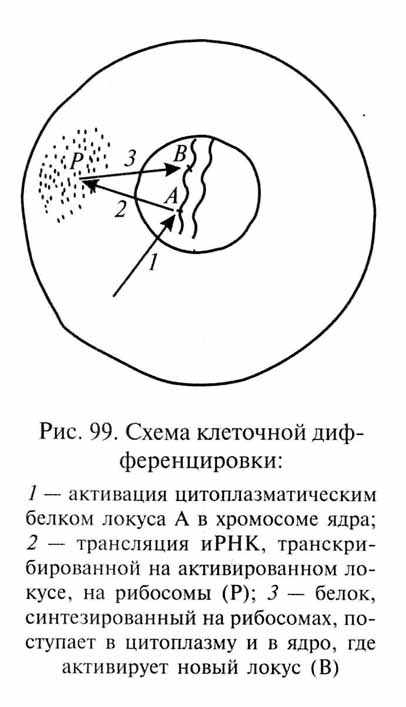
Очевидно, при эквивалентности генома всех зародышевых клеток их первоначальная дифференцировка должна определяться разнокачественностью их цитоплазмы. В период дробления практически не происходит роста клеток между их последовательными делениями. Поэтому в разные бластомеры попадают различные и, вероятно, разнокачественные участки цитоплазмы зиготы-Возможно, разнокачественность цитоплазмы и определяет активацию различных генов в разных бластомерах. Деятельность активированных генов приводит к изменениям цитоплазмы бластоме-ров - специфическим в разных клетках. Измененное цитоплазма-тическое окружение взаимодействует с геномом клетки, вызывая избирательную активацию новых групп генов и репрессируя другие группы. Так запускается цепь последовательных взаимодействий между геномом и цитоплазмой, протекающая по принципу обратной связи (положительной для одних и отрицательной для других генных комплексов), в результате которых клетка дифференцируется в определенном направлении (рис. 99). Однако пока еще мало известно о конкретных взаимодействиях между ядром и цитоплазмой на разных этапах клеточной дифференцировки.
Онтогенез отнюдь не сводится к дифференциации разных типов клеток. Важнейшим аспектом индивидуального развития многоклеточных организмов является установление общего плана их строения, т. е. оформление пространственной структуры организма и определенного расположения его частей. У всех более сложных многоклеточных организмов к этому добавляются процессы органогенезов - формирование дифференцированных органов. Все ^эти процессы осуществляются посредством взаимодействия друг с другом отдельных клеток и клеточных комплексов.
О сущности этих взаимодействий известно, пожалуй, еще меньше, чем о молекулярных механизмах клеточной дифференцировки, хотя исследования в этой области ведутся уже давно. Одной из Наиболее разработанных концепций является теория эмбриональной индукции, созданная трудами Г.Шпемана и его учеников.
Под эмбриональной индукцией понимается такое взаимодействие между двумя клеточными комплексами зародыша, при котором один из них - организатор или организационный центр - Направляет (индуцирует) дифференциацию другого - реагирующей ткани - в определенном направлении. В нормальных условиях, т. е. в живом зародыше, присутствие организатора необходимо Для формирования из реагирующих тканей специфических структур взрослого организма. Так, у зародышей позвоночных крыша первичной кишки, образованная зачатком хорды, индуцирует в расположенной над ней эктодерме развитие нервной системы; глазной бокал, растущий по направлению к покровам от зародышевого головного мозга, индуцирует развитие хрусталика глаза; зачаток внутреннего уха (слуховой пузырек) индуцирует развитие вокруг себя слуховой капсулы и т.п. Согласно теории эмбриональной индукции, дифференциация органов в индивидуальном развитии представляет собой цепь индукционных взаимодействий, в которую последовательно включаются новые и новые организаторы, и каждый предшествующий этап "запускает в действие" следующий.
Характер взаимодействий между морфогенетическими системами в процессах эмбриональной индукции может изменяться в филогенезе. Например, у эмбрионов ряда костных рыб (Amia, Salmo) невромасты сейсмосенсорных рецепторов (или органов боковой линии) в области головы играют роль индукторов для формирования накладных костей черепа. Однако у хрящевых рыб невромасты сейсмосенсорных рецепторов в области головы имеются, но накладные кости черепа не развиваются. Следовательно, индукционные взаимодействия здесь разорваны: либо невромасты потеряли свойства индукторов, либо реагирующие ткани утратили свою реактивность. В то же время у наземных позвоночных накладные кости черепа имеются, а органов боковой линии нет. Очевидно, в этом случае вместо старого механизма индукции возник новый. С другой стороны, некоторые индукционные системы поразительно консервативны, сохраняясь у очень далеких друг от друга групп - таков первичный хордомезодермальный организатор у всех хордовых.
Природа эмбриональной индукции еще не вполне выяснена. Индукция может основываться на непосредственном взаимодействии поверхностных мембран клеток организатора и реагирующей ткани, или на химической передаче индуцирующего действия, или на совокупности пространственных и химических взаимодействий. По некоторым данным, в химической индукции существенную роль может играть белковый компонент рибонуклеопротеи-
дов индуктора.
Явление эмбриональной индукции, несомненно, играет важную роль в онтогенезе, но отнюдь не исчерпывает всех форм взаимодействий между клеточными комплексами зародыша, обеспечивающих развитие структурной упорядоченности организма.
Недавно (80-90-е гг. XX в.) было показано, что важную роль в контроле процессов онтогенетического формирования пространственной организации тела зародыша играет особая категория генов-регуляторов, именуемых гомеобокссодержащими, или гомеобоксными. Гомеобокс представляет собой эволюционно консервативный генетический блок - последовательность из 180 нуклеотидов, обнаруженный в геноме филогенетически столь далеких друг от друга организмов, как позвоночные, иглокожие, насекомые и нематоды. Гомеобокс кодирует "гомеодомен" из 60 аминокислот. Протеины, содержащие этот гомеодомен, регулируют транскрипцию других генов. Гомеобоксные гены являются важными регуляторами, контролирующими онтогенетические программы, которые определяют формирование специфики разных сегментов вдоль передне-задней оси тела. Очевидно, мутации гомеобоксных генов могут иметь очень глубокие последствия для организма, приводя к изменениям соответствующих программ развития. Значительное большинство исследованных мутаций гомеобоксных генов (например, у мух-дрозофил) летальны, что вполне естественно для крупных мутаций. Эти обстоятельства определяют высокий эволюционный консерватизм гомеобоксных генов, сохраняющих значительную степень общности у филогенетически далеко разошедшихся групп организмов.
Проявление мутаций в фенотипе
Мутации проявляются фенотипически и подвергаются действию отбора, начиная с той стадии онтогенеза, когда мутантные гены оказываются активированными. Очевидно, общий фенотипический эффект данной мутации в среднем будет тем меньше, чем позднее в онтогенезе начинается функционирование мутантного гена. При этом фенотипический эффект мутации отнюдь не сводится к действию мутантного гена на признаки, им непосредственно контролируемые. Плейотропный (множественный) эффект влияния гена на фенотипические признаки складывается из взаимодействия различных морфогенетических систем в развивающемся организме, которые сами по себе могут иметь совершенно разную генетическую основу. Прекрасным примером этого может служить фенотипический эффект летальной мутации мышей, известной под названием "врожденная гидроцефалия". При этом у новорожденных мышей, погибающих от кровоизлияния в мозг, отечная лобная часть черепа заполнена геморрагической жидкостью; отсутствуют плоские накладные кости черепа; гипофиз имеет аномальную форму, выражены аномалии век и усов (вибрисс); морда укорочена. Сам же мутантный ген не влияет ни на один из этих признаков: он обусловливает лишь аномалии строения хрящевых клеток. Нарушения роста хрящевого эмбрионального черепа приводят к его укорочению, в результате которого мозг оттесняется кверху, что препятствует нормальному оттоку цереброспинальной жидкости. Это последнее приводит в конечном итоге ко всем перечисленным выше результатам (кровоизлияние в мозг, отсутствие плоских костей, аномалии век и др.). Мутация, обусловливающая сама по себе лишь аномалию хрящевых клеток, благодаря взаимодействиям различных зародышевых структур на эпигенетическом уровне, влечет за собой целую серию вторичных и третичных фенотипических эффектов, не относящихся непосредственно к компетенции мутантного гена.
Поскольку мутации со значительным фенотипическим эффектом (рано выявляющиеся в онтогенезе) имеют гораздо меньше шансов на сохранение, чем более поздно выявляющиеся малые мутации, онтогенез в целом приобретает тенденцию оставаться консервативным из поколения в поколение. В наибольшей степени это относится к основным процессам формообразования (установление общего плана строения, морфогенез важнейших органов), которые протекают чрезвычайно сходно у всех родственных видов в пределах очень крупных таксономических групп, таких, как классы и даже типы. С этим связано действие эмпирического обобщения, известного как "закон зародышевого сходства", сформулированного одним из крупнейших эмбриологов XIX в. К.Э. фон Бэром: "Развитие всех животных идет таким образом, что прежде всего определяется основной тип, причем гистологическая и морфологическая дифференцировки очень малы и только начинаются. Эмбрион при дальнейшем развитии этих дифференцировок переходит из своей основной формы в вариацию ее, т. е. из основной формы с характером данного класса переходит в таковую с характером отряда или семейства этого класса или в еще более ограниченную форму, пока наконец не появляются особенности индивидуума" (рис. 100).

Взаимоотношения онтогенеза и филогенеза
Проблему взаимоотношений индивидуального развития организмов и их эволюции затрагивал уже Ч.Дарвин, подчеркивавший влияние естественного отбора на все стадии индивидуального развития и формирование под контролем отбора специфических личиночных приспособлений. Позднее эту проблему разрабатывали Ф. Мюллер, Э. Геккель, В.Гарстанг, Г. де Вир и другие ученые.
В эволюционно-морфологическом плане соотношение онтогенеза и филогенеза наиболее полно было рассмотрено А.Н.Северцовым в его теории филэмбриогенезов (1912-1939). Согласно этой концепции филогенез не следует рассматривать как ряд сменянющих друг друга статических взрослых состояний организмов последовательных поколений (понимание филогенеза как эволюции взрослого состояния до сих пор очень распространено благодаря концентрации внимания ученых-неэмбриологов на взрослых организмах).
По А.Н.Северцову, филогенез представляет собой генетический ряд онтогенезов: эволюционируют не только фенотипы взрослых организмов, но и их онтогенезы, или кратко - филогенез есть эволюция онтогенеза. Изменчивость организмов проявляется на всех стадиях индивидуального развития и служит материалом для действия естественного отбора.
А.Н.Северцов разработал классификацию разных типов филогенетических изменений онтогенеза, которые он прежде всего разделил на ценогенезы и филэмбриогенезы. Ценогенезы представляют собой приспособления развивающегося организма к специфическим условиям эмбрионального или личиночного развития, которые не сохраняются во взрослом состоянии и не сказываются на особенностях последнего. Таковы, например, роговые зубы и жаберный фильтрационный аппарат личинок бесхвостых амфибий, плацента и зародышевые оболочки у эмбрионов млекопитающих и т.п. С другой стороны, филэмбриогенезы - это изменения онтогенеза, связанные с эволюционными преобразованиями каких-либо признаков взрослого организма. Филэмбриогенезы могут происходить как на самых поздних стадиях индивидуального развития (надставка, или анаболия), так и на средних (уклонение, или девиация) или на ранних (архаллаксис) стадиях. Из сказанного выше следует, что эволюция посредством анаболии в целом более обычна, чем посредством девиации и тем более архаллаксиса. При этом следует иметь в виду, что между разными формами филэмбриогенезов нет отчетливых границ. Более того, некоторые эволюционные изменения, первоначально возникшие как изменения поздних стадий онтогенеза (анаболии), могут в ходе дальнейшей эволюции сместиться на более ранние стадии, т. е. вторично преобразоваться в девиации и архаллаксисы. Это может быть связано с необходимостью более раннего выявления в онтогенезе нового адаптивного признака. Такой эффект достигается посредством более ранней онтогенетической активации соответствующих генов. Только на тех стадиях онтогенеза, которые не подверглись ценогенезам или филэмбриогенезам, сохраняется сходство с состоянием соответствующих стадий онтогенеза предков. Такое повторение предкового состояния - рекапитуляция - может сохраняться только для отдельных эмбриональных структур или процессов, но не для всего организма в целом, как полагал Э. Геккель. Обобщение Геккеля, известное как "основной биогенетический закон" (онтогенез есть сжатое и сокращенное повторение хода филогенеза), не имеет универсального характера и реально наблюдается лишь как частный, хотя и достаточно широко распространенный случай соотношений индивидуального и филогенетического развития - только для отдельных органов при условии их эволюционных преобразований по способу анаболии, т. е. изменения конечных стадий онтогенеза. Рекапитуляция целых стадий филогенетического развития для всего организма, которую постулировал "основной биогенетический закон" Э.Геккеля, крайне маловероятна.
В работах А.Н.Северцова и его учеников (Б.С.Матвеев, В.В.Васнецов, С.В.Емельянов и др.) были собраны многочисленные примеры различных форм эволюционных преобразований онтогенеза. Укажем лишь некоторые из них. По способу анаболии шла эволюция конечностей лошадиных (Equidae), что доказывается рекапитуляцией в онтогенезе конечностей современной лошади стадий Постепенной редукции боковых лучей конечности, весьма сходных с соответствующими филогенетическими стадиями (рис. 101). По способу анаболии, видимо, происходила и эволюция производных покровов у завропсид: ранние стадии морфогенеза пера птиц полностью соответствуют морфогенезу роговой чешуи рептилий, а последующие стадии развития пера как бы продолжают этот морфогенез дальше состояния, достигнутого у пресмыкающихся в роговых образованиях их кожи.

В качестве девиации А.Н.Северцов рассматривал преобразование брызгальца (древней жаберной щели, находившейся у рыбообразных предков наземных позвоночных между челюстной и подъязычной дугами висцерального скелета) в полость среднего уха тетрапод.
Примером архаллаксиса может служить значительное увеличение числа сегментов (сомитов) мезодермы (и соответственно их производных - позвонков, мышечных миомеров и т.п.) у змей по сравнению с их предками - ящерицами. Сегментация мезодермы происходит на ранних стадиях онтогенеза, почти одновременно с ее обособлением как зародышевого листка (стадия нейруляции).
Наконец, примеры ценогенезов назвал автор этого последнего термина Э. Геккель: это развитие амниона, аллантоиса, хориона и других эмбриональных и личиночных органов. Нужно при этом отметить, что Геккель понимал термин "ценогенез" гораздо шире, чем А.Н.Северцов: для Геккеля ценогенезами были любые изменения индивидуального развития, искажавшие ("фальсифицировавшие") консервативные признаки и процессы (или палингене-зы, по Геккелю), а не только провизорные эмбриональные органы, как понимал этот термин А.Н.Северцов. (Провизорные органы - структуры, существующие и функционирующие лишь в течение эмбрионального периода). В частности, в качестве ценогенезов Геккель рассматривал изменения темпов индивидуального развития различных органов, или гетерохронии. Как показали многочисленные более поздние работы (Ф.Кайбель, А.Н.Северцов, В.Гарстанг и др.), гетерохронии являются чрезвычайно распространенной формой эмбриональной изменчивости и предоставляют богатый материал для различных эволюционных преобразований - как ценогенезов, так и филэмбриогенезов. Гетерохронии могут изменять темпы эмбриональной дифферен-цировки различных органов и приводить к более ранней или более поздней их закладке в онтогенезе. Ускорение процесса эмбрионального развития какого-либо органа Геккель назвал акцелерацией, замедление - ретардацией.
Педоморфоз
Важное эволюционное значение может иметь педоморфоз - одна из форм эволюционных преобразований онтогенеза, основанная на гетерохрониях. Педоморфоз представляет собой генетически обусловленное (наследственное) сохранение во взрослом состоянии признаков более ранних стадий онтогенеза. Термин "педоморфоз" ("уподобление детенышу") был введен В.Гарстангом в 1922г.; его долгое время использовали преимущественно для обозначения сохранения во взрослом состоянии личиночных признаков. При наличии в жизненном цикле личиночной стадии, существенно отличающейся по строению, физиологии и образу жизни от взрослого организма, у некоторых видов в определенных условиях происходит задержка (ретардация) морфогенеза соматических структур при сохранении нормальных темпов развития половой системы. Личинки приобретают способность к размножению - половому (неотения) или партеногенетическому (педогенез). Эти явления представляют собой ненаследственные модификации онтогенеза: в других условиях личинки могут нормально метаморфизировать, превращаясь во взрослую стадию. Широко известным примером неотении являются аксолотли (рис. 102) - неотенические личинки американских саламандр - амбистом (Ambystoma tigrinum и др.). Неотения является результатом подавления активности эндокринных желез, стимулирующих метаморфоз (у земноводных - щитовидная железа). Воздействуя на аксолотлей соответствующими гормональными препаратами, можно добиться нормального метаморфоза - аксолотль превращается во взрослую амбистому.
При педоморфозе происходит наследственное изменение (уменьшение чувствительности соматических тканей личинки к гормонам, стимулирующим метаморфоз), что приводит к полной утрате взрослой стадии онтогенеза: жизненный цикл укорачивается, последней в нем становится стадия, которая была прежде личиночной. Такой педоморфоз произошел, например, в эволюции некоторых групп хвостатых земноводных (протеи, сирены, рис. 103). На основе педоморфоза сформировалась организация аппендикулярий (Appendiculariae) - одного из классов подтипа низших хордовых - оболочников (Tunicata). Вероятно, предки аппендикулярий имели жизненный цикл со свободноплавающей личинкой и прикрепленной неподвижной взрослой стадией, подобно современным асцидиям (Ascidiae). В результате педоморфоза у предков аппендикулярий прикрепленная взрослая стадия онтогенеза была утрачена, и строение современных представителей этой группы во многом остается близким к строению личинок асцидий (имеются хорда, сегментированные мышцы, спинной нервный ствол и др., рис. 104).

В последние десятилетия получило распространение широкое понимание термина "педоморфоз" как эволюционного изменения онтогенеза, связанного с сохранением во взрослом состоянии признаков любых более ранних стадий индивидуального развития, в том числе и неличиночных. При таком широком понимании термина, введенном С.Гулдом, педоморфоз включает также явление, которое описано Л.Больком в 1926 г. под названием "фетализация" - сохранение у взрослых организмов отдельных признаков эмбриональных стадий. Больк обратил внимание на наличие ряда фетализированных признаков у современного человека; к ним относятся, в частности, общие пропорции черепа с характерным куполообразным сводом и преобладанием мозговой коробки над челюстным аппаратом (см. рис. 94), а также некоторые другие особенности (форма ушной раковины, характер волосяного покрова). Среди этих признаков приспособительное значение имеет, вероятно, лишь фетализация общих пропорций черепа, которая создала у предков человека предпосылки, необходимые для вмещения и защиты головного мозга при прогрессировавшем увеличении его размеров. Фетализация других признаков могла развиться коррелятивно.
Некоторые аналогичные проявления фетализации наблюдаются у ряда пород домашних животных (собаки, кошки, рогатый скот), у которых в черепе, по сравнению с их дикими родственниками и предками, относительно большая мозговая коробка и относительно слабо развит челюстной аппарат. По наблюдениям К.Лоренца, для домашних животных характерно сохранение во взрослом состоянии определенной инфантильности поведения, благодаря которой эти животные признают "авторитет" человека как "патрона", "вожака стаи". Вероятно, в процессе искусственного отбора человек отбирал наиболее легко приручающихся животных, а таковыми были как раз те особи, которые сохраняли во взрослом состоянии черты инфантильности, т. е. обладавшие в некотором смысле "фетализированным поведением". Такое поведение должно основываться на соответствующих особенностях функционирования нервных центров. А фетализация последних может быть коррелятивно связана с указанными чертами строения черепа и других морфологических признаков, также приобретавших фетализированный облик. Так или иначе определенная степень фетализации поведения и ряда морфологических признаков характеризует многих домашних животных. Вероятно, отсутствие подобных особенностей поведения затрудняет приручение других диких животных, и в работе по созданию новых пород домашних животных следует учитывать необходимость отбора особей с наиболее инфантильным (фетализированным) поведением. Явления педоморфоза описаны и у растений: согласно А.Л.Taxтаджяну, у цветковых растений фетализированы многие органы спорофита (цветок, листья), а организация мужского и женского гаметофитов подверглась общему педоморфозу. (Нужно отметить, что А.Л.Тахтаджян, С.Гулд и ряд других ученых используют для обозначения педоморфоза или некоторых его форм термин "неотения", что приводит к путанице ненаследственных модификаций [неотения] и наследственных изменений онтогенеза [педоморфоз]).
Выделяют две основные формы педоморфных эволюционных изменений. Первая из них - акцелерационный педоморфоз, или прогенез, представляет собой результат ускоренного онтогенетического развития органов размножения, тогда как соматические структуры сохраняют прежние темпы морфогенеза. При этом организм в целом сохраняет ювенильный (или личиночный) облик, приобретая способность к размножению. После достижения половой зрелости обычно снижаются темпы морфогенеза и роста соматических структур, что приводит к обшей миниатюризации организма. Вероятно, таков был путь эволюции предков аппендикулярий. Возможно, посредством этого же эволюционного механизма были редуцированы мужской и женский гаметофиты у голосеменных и покрытосеменных растений. Вторая форма - ретардационный педоморфоз, или ретрогенез, представляет собой замедление темпов морфогенеза соматических систем организма при нормальном созревании половой системы и росте.
Вероятно, механизмы ретрогенеза могут быть различны на разных стадиях индивидуального развития в зависимости от факторов, контролирующих морфогенез на разных стадиях (например, при участии гормонов или еще до начала функционирования эндокринных желез). При этом особенно благоприятные условия для педоморфных преобразований создает личиночное развитие с метаморфозом, поскольку личинка представляет собой не только особую стадию онтогенеза, но и особый тип организации, отличающийся от взрослого организма и приспособленный к жизни в определенных условиях внешней среды, обычно в иных, чем взрослые особи того же вида. В связи с этим целесообразно различать три онтогенетических уровня ретрогенезов. При ретардации онтогенеза на эмбриональных стадиях происходит фетализация, приводящая к сохранению во взрослом состоянии эмбриональных признаков (как это было описано Л.Больком для человека). Еще один хорошо известный пример фетализации - задержка гистогенеза скелета на эмбриональных стадиях, что приводит к сохранению у взрослых позвоночных животных некоторых групп (круглоротые, хрящевые рыбы, в значительной степени современные хвостатые и бесхвостые амфибии) хряща как основной скелетной ткани. Задержка соматического онтогенеза у личинок (вплоть до полного подавления метаморфоза) приводит к ларвизации, или личиночному педоморфозу. При большей или меньшей степени ларвизации сформировалась организация таких групп хвостатых амфибий, как скрытожаберники (Cryptobranchidae), амфиумы (Amphiumidae), протеи (Proteidae) и сирены (Sirenidae). Наконец, морфогенез некоторых соматических органов может быть задержан уже на ранних постметаморфозных (или постнатальных) стадиях. В этом случае у взрослого организма сохраняются некоторые признаки ювенильных стадий онтогенеза - ювенилизация, проявления которой не столь ярки, как в двух других формах ретрогенезов, но зато наиболее обычны (например, некоторое облегчение скелета, редукция надглазничного валика, уменьшение размера зубов и другие изменения, происходившие при становлении организации неоантропов).
Основная эволюционная роль любого педоморфоза (и вообше гетерохронии) заключается в формировании новых комбинации фенотипических признаков, причем эти преобразования могут происходить без существенных генетических изменений, поскольку генетические механизмы возникновения гетерохронии сравнительно просты, основываясь на изменениях активности отдельных генов. Поэтому гетерохронии являются широко распространенной формой наследственной изменчивости онтоге-незов, предоставляющей обильный материал для действия естественного отбора.
Онтогенез и целостность организма
В предыдущих разделах мы рассматривали механизмы эволюционных преобразований онтогенеза. Однако необходимо принимать во внимание и противоположный аспект - обеспечение в онтогенезе целостности организма, с которой связана эволюционная устойчивость важнейших морфогенетических систем. Этот аспект онтогенеза также оказывает глубокое воздействие на характер эволюционных преобразований. Целостность организма обеспечивается разнообразными корреляциями, представляющими собой системы упорядоченных взаимодействий различных структур в онтогенезе. Коррелятивно связанными друг с другом могут быть вообще любые структурные и функциональные характеристики организма. Механизмы коррелятивных связей весьма различны. И.И.Шмальгаузен выделил три основных онтогенетических уровня корреляций.
1. Генетические корреляции основываются непосредственно на процессах, происходящих на уровне генома (биосинтез белков - например, при контроле нескольких оперонов общим геном-регулятором). В фенотипе генетические корреляции проявляются, например, в виде плеиотропного действия генов, т. е. влияния данного гена на различные признаки и свойства. Так, мутация "ко-роткоухость" у домовой мыши сопровождается укорочением нёба и уменьшением высоты морды; мутация "полидактилия" у кур приводит к одновременному удвоению первого пальца как на задних, так и на передних конечностях; мутация "белоглазие" у дрозофилы сказывается также на плодовитости мух (снижая ее) и продолжительности жизни (уменьшение) и т.п.
Некоторые генетические корреляции связаны с образованием внутри генотипа генных комплексов "супергенов", интегрированных различными способами. Один из таких способов - возникновение хромосомных мутаций типа инверсий, затрудняющих Кроссинговер в мейозе с хромосомой, имеющей "нормальную" Последовательность генов. Отдельные гены, входящие в состав Подобного супергена, могут контролировать развитие разных фе-Нотипических признаков. Отбор будет благоприятствовать сохранению таких супергенов, если объединяемые в них генные комплексы обеспечивают более высокую приспособляемость особей.
Наконец, как генетическую корреляцию можно рассматривать также эффект сцепления генов, локализованных в одной хромосоме поблизости друг от друга, хотя эта корреляция относительно легко нарушается кроссинговером и, вероятно, не имеет большого эволюционного значения.
2. Морфогенетические корреляции представляют собой результат взаимодействия различных зачатков в ходе эмбрионального развития. Примеры такого взаимодействия дают, в частности, явления эмбриональной индукции. Изменение индуктора влечет за собой соответствующие изменения индуцируемых структур. Если последние, в свою очередь, являются вторичными индукторами, цепь коррелятивных изменений продолжается далее (пример подобной цепи первичных и вторичных индукторов представляют соотношения глазного бокала, хрусталика и роговицы глаза).
3. Морфофункциональные, или эргонтические, корреляции являются результатом взаимодействий различных структур в морфо-функциональных системах организма в постэмбриональном периоде онтогенеза. Так, ослабление мышечной активности при вынужденной длительной неподвижности конечности приводит к уменьшению кровоснабжения соответствующих мускулов. Изменения в функционировании желез внутренней секреции сказываются на функциональном состоянии различных органов, что нередко сопровождается и структурными изменениями последних. Например, усиление секреции андрогенов корой надпочечников приводит к общей маскулинизации организма, т. е. проявлению и усилению мужских особенностей, даже у генетически женских индивидов: усиленный рост скелета, развитие мускулатуры, развитие некоторых мужских и редукция женских вторичных половых признаков. Все подобные взаимосвязи носят характер функциональных корреляций.
Выделение трех указанных категорий корреляций, в сущности, условно. Это ярко проявляется в рассмотренном выше плейо-тропном эффекте мутации гидроцефалии у мышей. Разнообразные патологические изменения головного мозга, черепа, глазных век, вибрисс и др. возникают на основе морфогенетических корреляций (взаимосвязи в ходе онтогенеза), сама же мутация непосредственно сказывается лишь на процессах формирования эмбрионального хрящевого скелета. Очевидно, в этом случае тесно связаны явления генетических и морфогенетических корреляций. В свою очередь, морфогенетические корреляции нередко переходят в корреляции функциональные. Так, недоразвитие у молодых животных некоторых скелетных мышц, основанное на генетических причинах или определяемое условиями развития, приводит к недоразвитию отростков и гребней на поверхности костей, служащих для прикрепления этих мышц (например, для височных мышц млекопитающих - венечные отростки нижней челюсти). Таким образом, три онтогенетических уровня корреляций тесно взаимосвязаны, представляя в целом общую систему интеграции организма в его индивидуальном развитии.
Изучение коррелятивных систем организмов имеет важное значение для понимания процессов эволюционных преобразований. Филогенетические изменения относительных размеров различных структур могут определяться постоянным соотношением скоростей их роста. Два признака могут изменяться в эволюции так, что направление и скорость изменений одного зависят от изменений другого. Например, по данным Р.Робба, в филогенезе лошадей сохранялось постоянное соотношение удлинения морды и всего черепа в целом, хотя абсолютные скорости этих процессов были различны. В результате изменение пропорций черепа в эволюции лошадей в сторону удлинения морды происходило, вероятно, не посредством специальных мутаций, ответственных именно за эти конкретные изменения относительной и абсолютной длины морды, а коррелятивно - благодаря общему увеличению размеров тела и головы в филогенезе лошадей при постоянном соотношении длины морды и длины черепа в целом. Таким образом, увеличение размеров тела коррелятивно сказывалось на соответствующем удлинении морды по отношению к общей длине черепа. Следовательно, в этом и других подобных случаях филогенетические изменения конкретного морфологического признака, которые констатируются палеонтологами, происходили без каких бы то ни было специально ответственных за эти изменения мутаций. По определению Симпсона, в подобных случаях признак эволюционирует не генетически, а лишь морфогенетически. Подобные ситуации необходимо учитывать и при определении темпов эволюционных изменений различных признаков.
Благодаря наличию корреляций отбор, благоприятствующий какому-либо признаку, будет автоматически способствовать сохранению и распространению в популяциях всего комплекса коррелятивно связанных признаков, которые сами по себе могут иметь как нулевую (нейтральные признаки), так и слабо отрицательную приспособительную ценность. Поэтому нельзя ограничиваться действием отбора на какой-либо один признак - эффект отбора всегда определяется по общей приспособительной ценности всех взаимосвязанных изменений. При искусственном отборе, который селекционеры ведут обычно по отдельным интересующим их признакам, в результате происходит общее снижение приспособленности организмов, вплоть до потери ими жизнеспособности вне специфических условий их содержания человеком.
Формирование систем корреляций, совершенствующих интеграцию организма, в филогенезе происходило параллельно с усложнением (дифференциацией) организации, являясь важной составной частью эволюционных преобразований по пути морфо-физиологического прогресса. Можно выделить два основных пути формирования коррелятивных связей между фенотипическими признаками. Первый из них -- возникновение корреляций как случайный побочный результат перестроек морфогенетических систем, обусловленных возникновением каких-либо мутаций. В этом случае корреляции признаков на уровне фенотипа первоначально не имеют приспособительного значения (как опи
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





