|
ГЛАВА 2. ЖИЗНЬ В ПАЛЕОЗОЙСКОЙ ЭРЕ
Итак, в раннем кембрии произошел взрыв формообразовательной эволюции, который в течение примерно 15 млн лет привел к появлению в палеонтологической летописи представителей почти всех известных типов организмов. В течение палеозойской эры, продолжавшейся свыше 300 млн лет, произошли значительные изменения физико-географических условий: рельефа суши и морского дна, общего соотношения площади материков и океанов, положения континентов, климата и многих других факторов. Эти изменения неизбежно должны были сказываться на развитии жизни.
Арена жизни в палеозое
На протяжении палеозоя море неоднократно наступало на континент, заливая опускавшиеся участки континентальных платформ - морские трансгрессии начала кембрия, первой половины ордовика, когда была затоплена наибольшая за весь фанерозой часть континентов, раннего силура, девона и карбона. Морские трансгрессии сменялись поднятием суши, сопровождавшимся горообразовательными процессами и морскими регрессиями (отступление морей) во второй половине кембрия, в конце ордовика, в самом начале и в конце силура, в середине карбона и в пермском периоде. Мощные горные хребты поднялись в конце силура в Скандинавии, Шотландии, Гренландии, на востоке Северной Америки и в ряде других районов земного шара (Каледонская фаза горной складчатости), в каменноугольном и пермском периодах - в Средней Европе (Арденны, Судеты, Гарц, Шварцвальд и др.), на Урале и в Средней Азии (Алай, Тянь-Шань, Ку-энь-Лунь), на севере Сибири, в Австралии и др. (Герцинская фаза складчатости).
Согласно классической теории, которая берет начало в работах Ч.Ляйеля, происходившие в геологической истории изменения границ суши и моря объясняются вертикальными смешениями участков земной коры. Однако ныне все большее признание получает концепция мобилизма -- горизонтальных перемещении континентальных блоков, или "дрейфа континентов", выдвинутая в начале XX в. А. Вегенером. Современный вариант этой концепции - теория тектоники литосферных плит, или "новая глобальная тектоника" - - основывается на глобальном синтезе результатов геолого-геофизических наблюдений и во многом отличается от взглядов Вегенера.
По современным геофизическим данным, астеносфера - верхний слой верхней мантии Земли, располагающийся на глубинах 60-250 км, - имеет пониженную вязкость. Предполагают, что в мантии существуют конвекционные потоки ее вещества, источником энергии для которых являются, вероятно, радиоактивный распад и гравитационная дифференциация вещества мантии. Движение этих потоков со скоростью несколько сантиметров в год происходит в горизонтальной плоскости в верхнем слое верхней мантии от областей подъема к областям опускания ее вещества. В это движение вовлекаются литосферные плиты, как бы плавающие в состоянии изостатического равновесия на поверхности астеносферы и служащие основанием для континентов. При столкновении континентальных плит их края деформируются с возникновением зон складчатости и проявлений магматизма. При столкновении континентальной и океанической плит последняя как бы подминается первой и растекается под ней в астеносфере. Концепция мобилизма в ее современном виде не свободна от противоречий и, вероятно, еще подвергнется существенным перестройкам. Однако уже и теперь она дает удовлетворительное объяснение целому ряду как геологических, так и биологических проблем. К последним относятся, в частности, характерные особенности географического распространения многих видов животных и растений, а также некоторые исторические изменения фауны и флоры.
Согласно представлениям тектоники плит положение и очертания материков и океанов в палеозое были совершенно иными, чем теперь. Палеогеографические реконструкции, выполненные различными учеными с использованием разных методов, довольно существенно различаются в деталях. Однако большинство авторов согласны, что в кембрии южные материки (Южная Америка, Африка, Австралия, Антарктида, а также Индия) были объединены в суперконтинент, называемый Гондваной (рис. 34). Гондвана располагалась главным образом в Южном полушарии, от полюса (находившегося в современной Северо-Западной Аф-Рике) до экватора. В основном в экваториальной зоне находились меньшие по размерам северные материки: Северо-Американский, Восточно-Европейский, Сибирский и микроконтиненты (Средне-Европейский, Казахстанский и др.). Между ними и Гондваной был океан.

В ордовикском периоде северные континенты начинают постепенно сближаться друг с другом. В силуре и девоне Северо-Амери-канский континент объединился с Восточно-Европейским, с образованием Евроамерики. Пояс Каледонской складчатости в Северо-Западной Европе и Восточной Северной Америке сформировался при столкновении этих континентальных плит. В раннем карбоне сближение континентов продолжалось. Этому соответствовало возникновение поясов Герцинской складчатости. Процесс сближения континентов завершился во второй половине палеозоя (поздний карбон - ранняя пермь) образованием гигантского суперконтинента Пангеи, протянувшегося от Южного полюса, который сместился к этому времени в Антарктиду, далеко в Северное полушарие. Северные материки образовали в составе Пангеи комплекс, получивший название Лавразии.
Перемещения континентальных плит, сопровождавшиеся изменениями рельефа суши и морского дна, обусловливали изменения направлений морских течений и атмосферной циркуляции. Сложные взаимодействия всех этих факторов определяли климатические изменения как местного, так и глобального характера. Согласно Ч.Бруксу, относительное увеличение площади суши в высоких широтах, особенно расположение полюса на суше, вызывает общее понижение температуры поверхности планеты, тогда как увеличение площади суши в низких широтах и расположение полюсов в океане - повышение температуры.
В первой половине палеозойской эры на Земле господствовал в целом теплый климат: средняя температура поверхности Земли была сравнительно высокой, а перепад температур между полюсами и экватором невелик, климатическая зональность была выражена относительно слабо. Теплый период в конце палеозоя, начиная со второй половины карбона, сменился холодным, с более низкой средней температурой, резко выраженной климатической зональностью и значительной разницей температур между полюсами и экватором. Возможно, это связано с расположением в это время центральной части Гондваны в районе Южного полюса. В позднем карбоне и ранней перми мощный ледниковый щит покрывал Антарктиду, Австралию, Индию, южные части Африки и Южной Америки - это было время великого оледенения Гондваны. В верхнепалеозойских отложениях всех этих районов широко распространены тиллиты - ископаемые остатки древних ледниковых морен. В первой половине палеозоя тиллиты имели гораздо более ограниченное распространение; по-видимому, оледенение в конце ордовика и девоне затрагивало лишь приполярные районы (Северо-Западная Африка в ордовике, Аргентина в силуре и девоне). Северный полюс в палеозое находился в океане.
Жизнь в палеозойских морях и пресных водоемах
Вероятно, в кембрийском периоде, как и в докембрии, основные проявления жизни были сосредоточены в морях. Однако организмы уже заселили все разнообразие доступных в море местообитаний, вплоть до прибрежного мелководья, и, возможно, проникли в пресные водоемы.
Водная флора была представлена различными водорослями, основные группы которых возникли еще в протерозое.
Теплые моря кембрийского периода были обильно населены разнообразными животными. Наиболее богата была донная фауна неглубоких морей, прибрежных отмелей, заливов и лагун (во время морских трансгрессий мелководные местообитания получают очень широкое распространение). На морском дне жили разнообразные прикрепленные животные: губки, кишечнополостные (одиночные и колониальные представители различных групп полипов), стебельчатые иглокожие, плеченогие и др. Большинство из них питалось различными микроорганизмами (простейшие, одноклеточные водоросли, бактерии и др.), которых они различными способами отцеживали из воды (фильтрационное питание). Некоторые колониальные организмы, обладавшие известковым
скелетом, возводили на дне моря рифы, подобно современным коралловым полипам. В раннем палеозое это были гидроидные полипы-строматопоры (подкласс Stromatoporoidea), древние коралловые полипы-табуляты (подкласс Tabulata), мшанки (тип Bryozoa) и археоциаты (тип Archaeocyatha). Археоциаты - группа примитивных прикрепленных животных, широко распространенных в раннем кембрии, но полностью вымерших к концу силура. Археоциаты имели известковый скелет в виде своеобразного кубка (рис. 35), стенки которого пронизывали многочисленные поры. Размеры таких кубков варьировали от нескольких миллиметров до 40 сантиметров. Снаружи кубок был одет мягкой оболочкой. Мягкие ткани внутри кубка, видимо, имели губчатое строение, будучи пронизаны системой полостей.

Ряд других групп прикрепленных морских животных, также переживших расцвет в палеозойской эре, сохранились до наших дней в лице немногих видов. Таковы, например, стебельчатые иглокожие (подкласс Pelma-tozoa), представленные в палеозое 7 классами, из которых до современности дожили лишь морские лилии (Crinoidea). Сходная судьба была и у плеченогих - своеобразных прикрепленных животных, снабженных двустворчатой раковиной и внешне напоминающих двустворчатых моллюсков, но резко отличающихся от них внутренним строением. Раковина плеченогих имеет спинную и брюшную створки, а не две боковые, как у моллюсков. Замечательно, что с кембрийского периода до наших дней просуществовал род брахиопод Lingula (см. рис. 17), который, судя по ископаемым остаткам, сохранил свое строение практически без изменений свыше 500 млн лет, став одним из наиболее ярких примеров эволюционного консерватизма.
С морским дном были тесно связаны и многие другие группы животных. Различные черви приспособились к роющей жизни в толще донных осадков. Среди них были примитивные вторичноротые - кишечнодыша-Щие (Enteropneusta), принадлежащие к типу полухордовых. Наиболее древние ископаемые остатки этих животных были обнаружены в среднекембрийских отложениях (фауна Бёр-Джес), но, несомненно, возникла эта группа еще в докембрии.
Среди водорослей и кораллов по морскому дну медленно передвигались свободноживущие иглокожие (представители подтипа Eleutherozoa - морские звезды, офиуры, голотурии и др.) и моллюски, основные группы которых известны, начиная с кембрия. Свободноживущие иглокожие возникли, вероятно, от прикрепленных предков, унаследовав от них внешнюю радиальную симметрию тела. Организация моллюсков сложилась в процессе приспособления древних червеобразных животных к питанию различными донными обрастаниями и остатками погибших организмов, пищей обильной и легкодоступной. Такой способ питания не требовал большой подвижности. А для защиты мягкого тела у моллюсков развилась прочная и тяжелая известковистая раковина, еще более ограничившая их подвижность.
Но одна из групп моллюсков, вопреки общей эволюционной тенденции этого типа к развитию малоподвижных донных форм, уже в кембрии сумела перейти к освоению свободного плавания. Это были первые головоногие -- наутилоидеи, или кораблики (Nautiloidea), дожившие в лице единственного рода Nautilus (рис. 36) до наших дней. Раковина корабликов преобразовалась в гидростатический аппарат, позволяющий этим моллюскам изменять свою плавучесть, поднимаясь или опускаясь в толще воды без мышечных усилий. Раковина разделена поперечными перегородками на камеры, тело животного помещается в последней, самой большой из них. Перегородки пронизаны сифоном - длинным выростом тела, в котором проходят кровеносные сосуды. Камеры раковины заполнены жидкостью, которая может всасываться в кровь и затем выводиться из организма почками; вес раковины уменьшается, и моллюск всплывает. При погружении происходят обратные процессы. Активное плавание головоногих в толще воды происходит посредством выброса струи воды из мантийной полости через специальную воронку - животное движется реактивным способом.

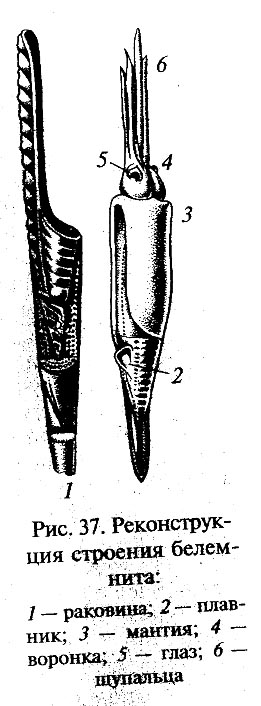
В девоне появились и другие группы головоногих моллюсков, в частности аммониты (Ammonoidea), имевшие, как и кораблики, спирально закрученную раковину. А в нижнем карбоне возникли первые представители высших головоногих, у которых раковина постепенно редуцировалась и оказалась заключенной внутри мягких тканей •тела (подкласс внутреннераковинных, Endocochlia). Эти преобразования были связаны с дальнейшей активизацией образа жизни головоногих. Древнейшими внутрен-нераковинными были белемниты (Belem-nitida), которые еще имели довольно большую удлиненно-коническую раковину (рис. 37), лишь прикрытую мантийной складкой тела1. В ископаемом состоянии наиболее часто сохраняется ростральная часть раковины белемнитов. Аммониты и белемниты играли видную роль в морской фауне верхнего палеозоя и мезозоя.
В толще и на поверхности воды в морях обитали и другие животные. Более примитивные группы использовали пассивное плавание, дрейфуя по течению и удерживаясь на поверхности с помощью особых плавательных пузырей или "поплавков", заполненных газом. Таковы плавающие колониальные кишечнополостные сифонофоры (Siphonophora), живущие в морях и поныне.Впервой половине палеозоя подобный образ жизни вели граптолиты (подтип Graptolithina типа полухордовых, рис. 38). Среди граптолитов были также "псевдопланктонные" виды, не имевшие поплавков и прикреплявшиеся к плавающим водорослям. Были и формы, прикреплявшиеся к морскому дну. Граптолиты, появившиеся в кембрии, полностью вымерли в раннем карбоне (большинство их исчезли еще до конца силура).


Уже в раннем кембрии существовали и столь высокоорганизованные животные, как членистоногие, представленные тремя обособленными подтипами: жабродышащими (Вгап-chiata), хелицеровыми (Chelicerata) и трилобитами (Trilobita). Последний подтип - трилобиты (рис. 39) - чисто палеозойская группа, достигшая расцвета в раннем кембрии, когда они составляли до 60% всей фауны. Трилобиты сохраняли свои позиции до середины ордовика, после чего началось их угасание. Они окончательно вымерли к середине пермского периода. Для трилобитов характерно расчленение защищавшего тело спинного щита двумя продольными бороздами на 3 доли (отсюда и название подтипа). Эти членистоногие были чрезвычайно разнообразны. Их размеры варьировали от 1 до 75 см, хотя большинство не превышало 10 см. Трилобиты были в основном придонными животными. Они ползали по дну на членистых конечностях, состоявших каждая из двух ветвей, но могли, вероятно, и плавать в толще воды (среди трилобитов были даже формы, специализированные к плаванию), и зарываться в песок. По способу питания большинство трилобитов были, по-видимому, микрофагами, отцеживавшими из воды мелкий планктон или поедавшими ил.
Среди водных палеозойских членистоногих были и хищники, из которых особенно выделяются эвриптериды (отряд Eurypterida, принадлежащий к подтипу хелицеровых), родственные современным скорпионам, но достигавшие длины около 2 м (рис. 40). Это были самые крупные членистоногие, когда-либо существовавшие на Земле. Эвриптериды известны, начиная с ордовика. Они достигли наибольшего расцвета в силуре и первой половине девона, но уже с середины девона число их видов начинает сокращаться. В нижней перми эти крупные хелицеровые исчезают. Возможно, судьба эвриптерид определилась тем, что примерно с середины девона в роли крупных водных хищников с ними начинают конкурировать различные группы хищных рыб. Позвоночные (подтип Vertebrata типа хордовых) оказались лучше приспособленными к быстрому плаванию в толще воды. Проблема передвижения у древних позвоночных была решена принципиально иначе, чем у членистоногих, которые плавали посредством гребных движений конечностей. У позвоночных развитие внутреннего осевого скеле-

|та (хорды, а затем позвоночника) позволило использовать практически всю мускулатуру тела для создания силы поступательного движения посредством волнообразных изгибаний туловища и мощных ударов основного движителя - хвостового плавника. После развития челюстей (см. стр. 130-131) позвоночные получили преимущество перед крупными членистоногими и в приспособлениях нападения и защиты: костные или хрящевые челюсти позвоночных, расположенные в толще приводящих их в движение мышц, механически прочнее и надежнее, чем наружные хитиновые челюсти членистоногих, возникшие из членистых конечностей.
Благодаря этим преимуществам позвоночные уже в девоне заняли доминирующее положение среди активно плавающих (нектонных) животных с крупными размерами тела. Членистоногие же оказались лучше приспособленными среди относительно мелких организмов: громадное большинство членистоногих ныне имеют небольшие размеры. Зато в этой сфере членистоногие развили ни с чем не сравнимое разнообразие мелких форм.
Позвоночные (Vertebrata) вместе с бесчерепными (Acrania) и оболочниками (Tunicata) входят в состав типа хордовых (Chordata). Позвоночные появляются в палеонтологической летописи, начиная с нижнего ордовика. Более древние хордовые животные, которые были общими предками позвоночных, бесчерепных и оболочников, до самого последнего времени оставались в ископаемом состоянии неизвестными. В связи с этим о происхождении хордовых выдвигалось много различных гипотез (вплоть до совершенно невероятных предположений о возникновении этих животных от головоногих моллюсков или от членистоногих, общий план строения которых принципиально отличается от такового хордовых). Выше мы уже рассматривали происхождение всех вторичноротых животных, вероятными общими предками которых были олигомерные черви, близкие по своей организации к современным кишечнодышащим полухордовым (Enteropneusta). Если эволюция полухордовых и иглокожих была связана с развитием приспособлений к жизни на морском дне (прикрепленной или роющей в толще донных осадков), древние хордовые, вероятно, перешли к активному плаванию в толще воды. Приспособления к нектонно-му образу жизни и создали предпосылки для дальнейшей прогрессивной эволюции этой группы. При активизации плавания отбор благоприятствовал усовершенствованию нервной системы, органов чувств и локомоторного аппарата. С этим связано развитие метамерии мышц тела и возникновение хорды - уникального внутреннего скелета. Хорда представляет собой продольный опорный тяж, прочность которого основана на несжимаемости жидкого содержимого особых вакуолей (принцип гидроскелета). Кроме того, в хорде имеются сократимые волокна, подобные мышечным; их сокращение повышает жесткость данного участка хорды и способствует быстрому выпрямлению соответствующего сегмента тела.
А.Н.Северцов сопоставил особенности организации разных подтипов хордовых, выбирая общие для всех групп признаки, которые можно истолковать как примитивные, и обрисовал гипотетический облик древнейших хордовых животных, потомков олигомерных червей и предков позвоночных, бесчерепных и оболочников. Этих гипотетических протохордовых А. Н.Северцов назвал примитивными бесчерепными (Acrania primitiva). Примитивные бесчерепные (рис. 41) обладали билатерально симметричным, сжатым с боков телом без обособленной головы и снабженным спинным и хвостовым плавниками. Скелет представлен хордой и соединительнотканными структурами. Пищеварительная система, как у всех низших хордовых, начиналась большой глоткой с эндостилем, наджаберной бороздой и жаберными щелями, пронизывающими ее боковые стенки; сзади глотка продолжалась коротким кишечником с печеночным выростом. Питание осуществлялось путем слизевой фильтрации: отцеживание из воды, прогоняемой мерцательным эпителием через глотку и жаберные щели, взвешенных в ней микроорганизмов и частиц ила, слизевым фильтром на жаберных щелях. Над хордой тянулась продольная нервная трубка - центральная нервная система. Кровеносная система включала продольные сосуды-аорты: брюшную (под глоткой) и спинную (под хордой), соединенные жаберными артериями. Мышцы всего тела имели метамерное строение.
В 1995-1999 гг. китайские палеонтологи Ж.Я.Чен, Г.К.Чжоу, Ч.В.Ли и др. сделали ряд замечательных находок ископаемых остатков древнейших хордовых животных в нижнекембрийских отложениях Юнънаня (Южный Китай). В тонкослоистых глинистых сланцах смогли сохраниться отпечатки структур, образованных мягкими тканями. Строение этих раннекембрийских хордовых. Названных юннанозооном (Yunnanozoon) и хайкоуэллой (Haiko-uella), оказалось очень близким к организации гипотетических Примитивных бесчерепных, обоснованной А. Н.Северцовым. Хай-Коуэлла, которая изучена более полно, была небольшим (25-40 мм) животным с ланцетовидным, сжатым с боков телом, дифференцированным на голову, туловище и короткий хвост. Имелись спинной, хвостовой и брюшной непарные плавники. Вдоль всего тела тянулась хорда. Мышцы тела были разделены на сегменты и сильнее развиты в спинной части туловища. На нижней стороне головы располагался окруженный небольшими щупальцами рот, который вел в объемистую глотку. По ее дну тянулся железистый желобок-эндостиль, а боковые стенки были пронизаны 6-7 парами жаберных щелей и укреплены жаберными дугами, на которых находились жаберные нити. Над хордой тянулся продольный тяж центральной нервной системы, который у хайкоуэллы в голове был дифференцирован как головной мозг с 3 отделами. Возможно, у хайкоуэллы имелись и парные глаза. Позади глотки находилось сердце, от которого кпереди тянулась под глоткой брюшная аорта, связанная жаберными артериями со спинной аортой.
Вероятно, расхождение филогенетических стволов ранних хордовых, которые дали начало подтипам позвоночных, бесчерепных и оболочников, произошло в самом раннем кембрии. Возможно, юннанозоон был близок к предкам бесчерепных, а хайкоуэлла - к предкам позвоночных. Ископаемые остатки древнейших представителей оболочников (Palaeobotridium) обнаружены в верхнекембрийских отложениях Невады в Северной Америке.
С позднего кембрия до позднего триаса существовала загадочная группа древних животных - конодонты, или конодонтофо-риды (Conodontophorida), долгое время известные лишь по многочисленным микроскопическим конусовидным зубам (которые, собственно, и именуются конодонтами), укрепленным небольшими группами или поодиночке в зубных пластинках. В 1983 г. в нижнем карбоне Шотландии были обнаружены ископаемые остатки мягкого тела животного (Clydagnathus), оотовой аппарат которого образован конодонтами. Это существо имело червеобразное уплощенное с боков тело длиной около 5 см, пару больших глаз, хвостовой плавник и метамерные мышцы. Электронно-микроскопические исследования показали, что конодонты образованы тканями, характерными для позвоночных животных, - хрящом, костью и эмалеподобным веществом. Д. Бриггс, Р.Олдридж и ряд других палеонтологов считают, что конодон-тофориды принадлежали к низшим хордовым и близкородственны с позвоночными или даже входят в состав последних.
Древнейшие позвоночные были рыбообразными животными, лишенными челюстей, с телом, защищенным панцирем (рис. 42). Отсюда название этой группы - - панцирные бесчелюстные (Ostracodermi). Их панцирь был образован крупными щитами и мелкими щитками и пластинками, состоящими из 3 слоев примитивных костных тканей (в том числе аспидина и дентина). Ас-пидиновые щитки были снаружи покрыты своеобразной шагренью из мелких зубчиков, образованных дентином и, вероятно, эмалью (подобной зубной). Голову этих животных сверху и сбоку, а иногда и снизу защищал сплошной головной щит с отверстиями для глаз, непарного органа обоняния и так называемого "теменного глаза" - светочувствительного верхнего придатка промежуточного мозга, игравшего у примитивных позвоночных важную роль в регуляции различных функций организма в соответствии с суточным и годовым циклами и с уровнем освещенности внешней среды. Задняя часть тела была покрыта небольшими щитками, не препятствовавшими изгибанию этого отдела и работе хвоста. Настоящих парных конечностей у остракодерм, видимо, еще не было, но по бокам тела позади головы имелась пара покрытых чешуями придатков или плавниковых складок.
Наличие у некоторых остракодерм хорошо окостеневшего черепа позволило выяснить ряд важных особенностей внутреннего строения этих животных. Оказалось, что многие признаки остракодерм Поразительно сходны с соответствующими особенностями современных миног и миксин, принадлежащих к классу круглоротых (Cyclostomata). Помимо отсутствия челюстей у остракодерм и круглоротых сходно строение жаберного аппарата (жаберные мешки, Расположенные внутри решетки скелетных висцеральных дуг), непарного органа обоняния, внутреннего уха (лишь с 2 полукружными каналами), головного мозга и др. Однако круглоротые, в отличие от своих палеозойских родственников, совершенно лишены костной ткани (их скелет представлен лишь хордой и хрящами). В ископаемом состоянии круглоротые почти не известны; лишь недавно в среднекарбоновых отложениях Иллинойса в США были открыты ископаемые остатки миног (Mayomyzon) и мик-син (Myxinikela). Может быть, к предкам круглоротых близок Palaeospondylus - загадочная среднедевонская форма с хорошо сохранившимся позвоночником, черепом, напоминавшим череп круглоротых, и хвостовым плавником, но без всяких признаков костного панциря.
Современные круглоротые являются полупаразитами (миноги) и трупоедами (миксины). Остракодермы, вероятно, питались илом или отцеживали из воды микропланктон с помощью слизевого фильтра на жаберных щелях, как и личинки современных миног. У некоторых остракодерм обнаружены следы структуры, сходной с эндостилем - железистым желобком на дне глотки, клетки которого выделяют слизь для слизевой фильтрации.
Наличие хорошо окостеневшего скелета у раннепалеозойских остракодерм при отсутствии окостенений у круглоротых привело к тому, что мнения исследователей о соотношениях скелетных тканей в эволюции и онтогенезе позвоночных разделились. А.Н.Северцов, Д.В.Обручев и ряд других ученых аргументировали теорию, согласно которой древнейшие позвоночные не имели костного скелета и поэтому не сохранились в ископаемом состоянии. Костный скелет начал формироваться в покровах в виде отдельных "кожных зубов", подобных мелким плакоидным чешуям современных хрящевых рыб - акул и скатов. В дальнейшем происходило срастание этих чешуи друг с другом с образованием все более массивных комплексов. Из верхнесилурийских и девонских отложений известны остатки остракодерм (Thelodus; см. рис. 42), лишенных тяжелого костного панциря, но покрытых плакоидными чешуйками.
Однако все более древние остракодермы (из ордовика и раннего силура) имели хорошо развитый костный скелет, причем плакоидные чешуйки образовывали наружный слой щитков панциря. В связи с этим палеонтологи А.Ш.Ромер, Л.Хальстед и др. считают более вероятным, что элементы костного скелета появились у древнейших позвоночных по крайней мере столь же давно, как и хрящевые структуры. В сущности, хрящ и кость представляют собой лишь крайние состояния скелетных тканей позвоночных, связанные промежуточными формами. Хрящ более пластичен, менее прочен, чем кость, и способен как к поверхностному, так и внутреннему росту. Благодаря этому хрящ является лучшей эмбриональной скелетной тканью, а кость - лучшей основой скелета взрослых позвоночных.
В филогенезе позвоночных происходила активизация образа жизни: переход от придонной жизни к нектонной, от бенто- и планктонофагии к хищничеству. При этом отбор благоприятствовал освобождению тела от тяжкого бремени защитного костного панциря, характерного для ранних остракодерм. Облегчение тела происходило по-разному: либо путем постепенного преобразования крупных щитов в гибкий и легкий чешуйный покров, либо посредством редукции костной основы - с сохранением поверхностных плакоидных чешуи (телодонты, хрящевые рыбы) или с полной утратой и этих последних (круглоротые). Второй путь преобразований (полная или частичная утрата костной ткани) был, очевидно, проще по механизму перестроек онтогенеза (с задержкой развития скелета на эмбриональной стадии) и мог поэтому осуществиться быстрее, чем более сложное постепенное преобразование скелетных структур, требовавшее существенных перестроек морфогенетических механизмов.
В филогенезе позвоночных были испробованы разнообразные варианты перестроек примитивного тяжелого скелета; судьба разных эволюционных ветвей при этом сложилась различно. В конечном итоге наибольший успех выпал на долю тех групп позвоночных, которые избрали более медленный, но гораздо более перспективный путь усовершенствования костного скелета, а не отказались от него полностью. Поэтому утрату костного скелета у таких групп, как круглоротые или хрящевые рыбы, можно с полным основанием рассматривать как пример инадаптивной эволюции, как ее понимал В. О. Ковалевский.
Остракодермы пережили свой расцвет в силуре и первой половине девона. К концу девона панцирные бесчелюстные вымирают, вытесненные более прогрессивными группами позвоночных, принадлежащими к стволу челюстноротых (Gnathostomata). Челюстноротые появились в палеонтологической летописи в позднем силуре (остатки древнейших рыб из группы акантодий - Acanthodii); в девоне существовали уже разнообразные группы всех классов рыб. Их общие предки пока остаются неизвестными. Некоторые палеонтологи допускают возможность происхождения челюстноротых от примитивных остракодерм из группы разнощитковых (Heterostraci), которые не имели таких специфических особенностей бесчелюстных, как непарный орган обоняния, открывающийся на верхней стороне головы, энтодермальные жаберные мешки и др. Расхождение филогенетических стволов Позвоночных, ведущих к бесчелюстным (классы остракодерм и Круглоротых) и челюстноротым (все остальные классы позвоночных) произошло, вероятно, в позднем ордовике или раннем силуре. Это расхождение было связано с различными путями интенсификации жаберного дыхания, рассмотренными нами выше. Как мы видели, преобразования висцерального скелета на пути, избранном челюстноротыми, привели к формированию челюстного аппарата, наличие которого вместе с развитием парных конечностей стало важнейшим преимуществом, обеспечившим победу челюстноротых в конкуренции с бесчелюстными. Современные круглоротые насчитывают всего лишь около 50 видов против примерно 20000 видов различных групп рыб.
Древнейшие представители челюстноротых, обитавшие в морях и пресных водоемах раннего и среднего девона, были, как и остракодермы, одеты более или менее развитым костным панцирем, особенно мощным у панцирных рыб -- плакодерм (класс Placodermi). Панцирь плакодерм (рис. 43) состоял из двух подвижно сочлененных друг с другом частей, одна из которых защищала голову, а другая - переднюю часть туловища. По бокам передней части туловища располагались подвижные придатки, возможно гомологичные передним конечностям других челюстноротых.

У плакодерм не было настоящих зубов, их заменяли острые бивнеобразные выступы челюстных костей (у других групп челюстноротых зубы возникли из плакоидных чешуи кожного панциря, переместившихся по краям рта на челюсти). Среди плакодерм были мелкие придонные формы, вероятно конкурировавшие с остра-кодермами, и крупные хищники, например верхнедевонская ди-нихтис (Dinichthys), у которой лишь одетая панцирем передняя часть тела имела длину около 3,2 м. Плакодермы вымерли в раннем карбоне.
Другой примитивной группой палеозойских челюстноротых были уже упоминавшиеся акантодии (Acanthodii). Их тело покрывали многочисленные костные пластинки и крупные чешуи, были окостенения и во внутреннем скелете. У акантодии еще не установилось обычное для позвоночных число пар конечностей; по бокам тела (рис. 44) тянулся ряд плавников (до 7 пар), возникших, вероятно, в результате дифференциации первоначальных плавниковых складок. В дальнейшем средние плавники, расположенные ближе к центру тяжести тела (и поэтому малоэффективные как рули глубины и стабилизаторы движения) подверглись ре-дукции, так что сохранились лишь передняя и задняя пары. Челюсти у акантодии были связаны с черепом без участия второй висцеральной дуги (подъязычной). Такое состояние также очень примитивно: у всех остальных рыб верхний (гиомандибулярный) элемент подъязычной дуги становится подвеском суставной области челюстной дуги. Акантодии обитали в пресных и морских во-Доемах с позднего силура до середины пермского периода, когда эта древняя группа подверглась вымиранию.

В первой половине девона существовали уже и представители трех подклассов костных рыб (класс Osteichthyes): лучеперых (Actinopterygii), двоякодышащих (Dipnoi) и кистеперых (Crosso-pterygii). Первая группа, эволюция которой была в основном связана с освоением всей толщи воды в пресных водоемах и морях, в палеозое была относительно малочисленна. Оба других подкласса, объединяемые в группу хоановых рыб (Choanichthyes), напротив, пережили расцвет в палеозое. Их организация сформировалась в процессе освоения широко распространенных в палеозое пресных водоемов типа обширных мелководных озер, лагун и эстуариев, хорошо прогреваемых солнцем, обильно заросших водной растительностью и отчасти заболоченных. В воде таких водоемов из-за гниения растений нередко возникал дефицит кислорода. Важнейшим приспособлением к жизни в таких водоемах были легкие как дополнительный орган дыхания, позволявший использовать кислород воздуха при его недостатке в воде. Легкие представляют собой слепые мешковидные выпячивания самой задней части глотки, куда рыба через рот заглатывает воздух, используя для этой цели обычный жаберный насос.
(Вероятно, орган, соответствующий легким, возник очень рано в эволюции костных рыб; его гомологом у актиноптеригий является плавательный пузырь, служащий гидростатическим аппаратом, но используемый примитивными лучеперыми и как легкое. По некоторым данным, этот орган мог возникнуть еще у примитивных челюстноротых - общих предков плакодерм, хрящевых и костных рыб.).
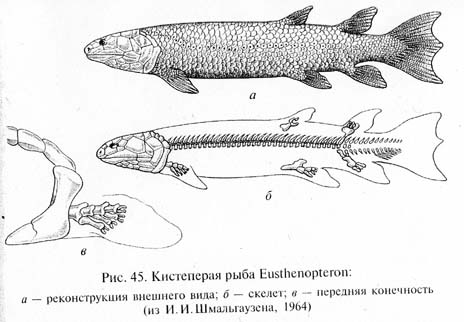
Хоановые рыбы вели в основном скрытный придонный образ жизни, охотясь в гуще растительности. Кистеперые были хищниками, подстерегавшими добычу в укрытиях, а двоякодышащие приспособились к питанию малоподвижными животными с твердым панцирем (моллюски, ракообразные). Плавники хоановых рыб приобрели особое строение, удобное для передвижения у дна водоемов: наружная "рулевая" лопасть, образованная тонкими плавниковыми лучами, редуцировалась, зато был сильно развит их внутренний костный скелет (базальные и радиальные элементы), выдвинутый из тела наружу вместе с мышцами, образуя мясистую лопасть в основании плавника (рис. 45). Такие плавники, называемые саркоптеригиями, позволяли рыбе переползать по дну, скрываясь среди растений.
Для ориентации в подводных зарослях важную роль приобретает обоняние. Чтобы обеспечить постоянный обмен воды в органе обоняния, задние ноздри (у рыб в каждый обонятельный мешок ведут два отверстия - переднее и заднее) перемещаются под верхнюю челюсть, становясь внутренними ноздрями, или хоанами (отсюда название "хоановые рыбы"). Хоаны позволяют вентилировать орган обоняния, используя изменения давления в рото-глоточной полости, которые возникают при движениях жаберных крышек в процессе вентиляции жабер.
Кистеперые рыбы были очень многочисленны в пресных и солоноватых водоемах в девоне и начале карбона, но сокращение их местообитаний при усилении горообразовательных процессов и общем поднятии материков постепенно привело эту группу к упадку. Одна из линий эволюции кистеперых рыб - целаканты (Coelacant-hiformes) - вторично перешли к жизни в море, и два вида этой группы сохранились до нашего времени в водах Мозамбикского п
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





