|
ГЛАВА 5. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Человек относится к отряду приматов (Primates), и эволюционная история человека есть часть филогенеза этой группы. Поэтому рассказ о происхождении человека целесообразно начать с анализа общих признаков приматов, среди которых можно найти предпосылки для развития особенностей строения, физиологии и поведения человека.
Приматов можно кратко охарактеризовать как группу лесных теплолюбивых плацентарных млекопитающих, приспособившихся к лазающему древесному образу жизни, сохранив достаточно примитивную основу организации. Примитивные черты в строении приматов проявляются в сохранении ими пятипалой конечности, опирающейся при ходьбе на всю ступню (плантиградность); в сохранении ключиц, которые утрачиваются при совершенствовании бега, в отсутствии специализации питания (большинство приматов всеядны и питаются как растительной, так и животной пищей) и т. д. Сохранение некоторых примитивных признаков и отсутствие узкой специализации соответствующих органов способствовали высокой эволюционной пластичности приматов, проявивших в ходе своего филогенеза способность адаптироваться к самым различным условиям существования и способам использования природных ресурсов.
Важнейшие особенности приматов связаны с развитием приспособлений к древесной лазающей жизни. Прежде всего это совершенствование хватательной способности конечностей, с которым связано приобретение возможности вращать кисть и предплечье вокруг их продольной оси (способность к пронации и супинации кисти, т.е. к ее вращению ладонью вниз и вверх), а также развитие противопоставления больших пальцев всем остальным, облегчающего хватание. Эти приспособления значительно увеличили общую манипулятивную способность конечностей, т. е. способность различным образом действовать с удерживаемым пальцами предметом.
Лазание по веткам требует хорошо развитого осязания - для исследования поверхности опоры. В связи с этим на концевых фалангах пальцев развились мясистые пальцевые подушечки, кожа которых богата осязательными тельцами и нервными окончаниями, а когти преобразовались в ногти с плоской и тонкой когтевой пластинкой, защищающей конец пальца лишь сверху.
Но самую важную роль среди органов чувств при лазании приобретают глаза, поскольку прыжки с ветви на ветку требуют объемного (стереоскопического) зрения с точной оценкой расстояний и надежности опоры. Развитие стереоскопического зрения достигается перемещением глаз на переднюю (лицевую) поверхность головы, их сближением и все большим перекрыванием полей зрения. Это, собственно, и создает бинокулярный эффект: предмет рассматривается двумя глазами с разных сторон, и восприятие обоих глаз интегрируется в центральной нервной системе.
С другой стороны, на деревьях падает роль обоняния. Соответственно орган обоняния у приматов подвергается некоторой редукции.
Чрезвычайно важные изменения, связанные с адаптацией к лазанию, произошли в центральной нервной системе. Передвижение по веткам представляет собой один из наиболее сложных способов локомоции, требующий точной оценки расстояний и качества опоры и соответствующей коррекции движений. На деревьях каждый шаг и прыжок представляет собой, по удачному выражению Я.Я.Рогинского, "отдельный творческий акт". Все это требует высокого совершенства функций центральной нервной системы - как в анализе непрерывно меняющейся обстановки, так и в осуществлении тонкого мышечного контроля разнообразнейших оттенков движений и положения тела. При совершенствовании всех указанных функций естественный отбор благоприятствовал прогрессивному развитию головного мозга, особенно коры больших полушарий, в которой формируется сложная система борозд и извилин (у низших приматов кора имеет гладкую поверхность)1, а также мозжечка. Увеличение относительного и абсолютного размера головного мозга возможно только при соответствующих изменениях пропорций черепа, в котором относительно увеличивалась мозговая коробка и уменьшался челюстной отдел.
Кочевая жизнь на деревьях, которую ведет большинство обезьян, не позволяет самкам иметь одновременно много детенышей, так как они, цепляясь за шерсть матери, сковывают ее движения. Поэтому отбор благоприятствовал тенденции к уменьшению числа детенышей у приматов до 1-2 на самку за одну беременность. Для компенсации малочисленности потомства у обезьян сформировались сложные формы родительского поведения, обеспечивающие длительный уход за детенышами и защиту их родителями. Результатом уменьшения числа детенышей была и редукция числа млечных желез до единственной грудной пары.
Очень важной характеристикой приматов является их общественный (стайный) образ жизни. Эффективная организация жизни стаи требовала развития особых (социальных) форм поведения. Наиболее жизнеспособными оказывались те стаи, в которых наследственно закреплялись такие формы поведения, которые обеспечивали снижение взаимной агрессивности и помощь более слабым животным в своей стае. В стаях формировалась так называемая социальная иерархия, т. е. происходило выделение разных категорий особей: доминантов - вожаков, животных "второго ранга" - субдоминантов - и последующих уровней иерархии, что отвечало задаче оптимальной организации жизни стаи: совместного целенаправленного движения по охотничьим угодьям, сведения к минимуму конфликтов между разными особями, преодоления различных критических ситуаций. Усложнение поведения в результате общественной жизни требовало, с одной стороны, дальнейшего совершенствования головного мозга, с другой - развития системы сигналов, которая позволила бы передавать необходимую информацию между разными членами стаи.
Таким образом, давление отбора в направлении совершенствования высшей нервной деятельности, ориентации в пространстве, Манипулятивных способностей конечностей и социального поведения, т. е. в конечном итоге в направлении формирования важнейших особенностей человека, определялось спецификой организации и образа жизни приматов.
Ранние этапы эволюции приматов
Приматы возникли от примитивных насекомоядных млекопитающих (Insectivora), отдельные группы которых перешли к лазающей древесной жизни. К основанию эволюционного ствола приматов, возможно, близки древесные землеройки - тупайи (Tupaiidae), обитающие ныне в тропических лесах Малайского полуострова и Филиппинских островов.
Эволюционная ветвь насекомоядных, ведущая к приматам, обособилась, вероятно, еще до конца мезозоя. Из позднемело-вых отложений известны ископаемые остатки животных, которых ряд палеонтологов рассматривает уже в качестве примитивных приматов. Это были пургатории (Purgatorius) - мелкие зверьки с признаками приспособлений к лазающей древесной жизни. Вероятно, они имели ночную активность, как и другие мезозойские млекопитающие, и питались насекомыми и плодами деревьев.
Из самых нижних горизонтов палеоцена известны остатки лемуров (Lemuroidea) - наиболее примитивной группы среди современных приматов. Лемуры ныне обитают в тропических лесах Мадагаскара, Африки и Южной Азии. Они ведут сумеречный и ночной древесный образ жизни. Это небольшие животные, в среднем достигающие размеров кошки, с длинным хвостом, пушистым мехом и удлиненной мордочкой. Глаза у лемуров обращены еще не прямо вперед, как у более высокоразвитых приматов, а несколько косо - вперед и вбок. У лемуров сохраняется довольно хорошо развитое обоняние. Этих симпатичных зверьков иногда называют "полуобезьянами", подчеркивая, что они еще не приобрели всего комплекса признаков, характерного для высших приматов, и занимают по уровню организации промежуточное положение между последними и древесными землеройками.
От каких-то примитивных лемуров в первой половине палеогена возникли настоящие обезьяны - антропоиды (Anthropoidea). Вероятно, их обособление от лемуров было связано с переходом к дневной активности, сопровождавшимся усилением роли зрения, увеличением размеров и совершенствованием строения головного мозга, развитием стайного образа жизни и связанных с ним социальных форм поведения.
По мнению ряда антропологов, от общего корня с антропоидами произошли тарзиевые, или долгопяты (Tarsioidea), - своеобразная обособленная группа ночных насекомоядных приматов, в организации которых причудливо сочетаются примитивные И специализированные черты.
Древнейшие (фрагментарные) остатки антропоидов известны из среднеэоценовых отложений Алжира (Algeripithecus) верхнеэоценовых слоев Бирмы и Египта. В раннем олигоцене существовали уже достаточно разнообразные их представители. Наиболее известен из них парапитек (Parapithecus), близкий по уровню организации к низшим мартышкообразным обезьянам (Cercopithecidae) и, возможно, принадлежавший к группе, которая была предковой для более высокоразвитых антропоидов.
До сих пор остается нерешенной проблема происхождения широконосых обезьян (Platyrrhini) Нового Света, сохранивших в целом более примитивное строение, чем современные обезьяны Старого Света, объединяемые в группу узконосых (Catarrhini). Названия "широконосые" и "узконосые" связаны с характерными различиями этих групп по ширине перегородки между наружными отверстиями ноздрей. Возможно, широконосые обезьяны возникли от раннеолигоценовых парапитековых в Африке и проникли в Южную Америку уже после ее обособления от Северной. Древнейшие ископаемые остатки обезьян в Южной Америке (Branicella) известны из олигоценовых отложений, образовавшихся на несколько миллионов лет позднее, чем нижне-олигоценовые отложения Египта с остатками парапитека. Однако согласно другой гипотезе, предковый ствол широконосых обезьян мог обособиться от основного ствола антропоидов еще в эоцене вместе с изоляцией Южной Америки, и дальнейшая эволюция широконосых и узконосых обезьян шла независимо и параллельно.
Из нижнеолигоценовых отложений Египта известны кроме парапитеков ископаемые остатки более высокоразвитых обезьян, названных проплиопитеками (Propliopithecus). Проплиопитек, судя по особенностям строения его нижней челюсти и зубов, мог быть предком гиббонов (Hylobatidae), которых многие ученые рассматривают уже как низших представителей человекообразных, или антропоморфных приматов (Hominoidea, или Anthropomorpha). Помимо гиббонов к антропоморфным приматам относят семейства высших человекообразных обезьян (Pongidae, или Simiidae), к которым принадлежат современные шимпанзе (Pan), гориллы (Gorilla) и орангутаны (Pongo), и семейство людей (Hominidae).
Антропоморфным приматам присущи наиболее крупные в пределах отряда размеры тела (самцы гориллы при росте около 180 см достигают веса 200 кг) и характерные пропорции: относительно короткое туловище и длинные конечности. У человекообразных обезьян передние конечности заметно длиннее задних. Вероятно, Пропорции тела и полувыпрямленная осанка антропоморфов сложились в связи с особыми формами передвижения этих крупных обезьян по деревьям, которые получили названия "круриация" и "брахиация". При круриации обезьяна идет на задних конечностях по нижним толстым ветвям деревьев в вертикальном или близком к нему положении, охватывая стопами опорную ветвь и придерживаясь передними конечностями за верхние ветви. Брахиация представляет собой более специализированную форму передвижения: это своего рода быстрый "полет" под кронами деревьев с перехватыванием руками за ветви и свисающие лианы, подобно прыжкам гимнастов на трапеции. Брахиация особенно характерна для гиббонов, но в какой-то мере как вспомогательный способ передвижения могла использоваться и общими предками всех антропоморфов. Во всяком случае, использование круриации и в некоторой степени брахиации древнейшими антропоморфами позволяет объяснить некоторые особенности их строения1.
Антропоморфы обладают наиболее сложными формами поведения, в том числе и социального, среди всех приматов, чрезвычайно высокой способностью к разнообразным манипуляциям с предметами, вплоть до изготовления примитивных орудий, т.е. предварительной обработки какого-либо объекта, которая делает его пригодным для осуществления заранее задуманного действия (например, обработка ветви руками и зубами для последующего добывания с ее помощью плода). Однако человекообразные обезьяны не обнаруживают способности к настоящей трудовой деятельности, принципиальной особенностью которой является изготовление орудий не с помощью естественных органов, а посредством других предметов, которые используются как искусственные средства обработки. С другой стороны, человекообразные обезьяны весьма способны к научению: наблюдения ряда этологов в природных условиях показали, что в стаях шимпанзе происходит своего рода обмен опытом между разными обезьянами путем наблюдения и перенимания операций, "изобретенных" более старыми и опытными животными. Однако даже у высших обезьян в отличие от человека не обнаружено способности к продолжительной концентрации внимания, требующей подавления посторонних отвлекающих впечатлений и необходимой для перехода к настоящей трудовой деятельности.
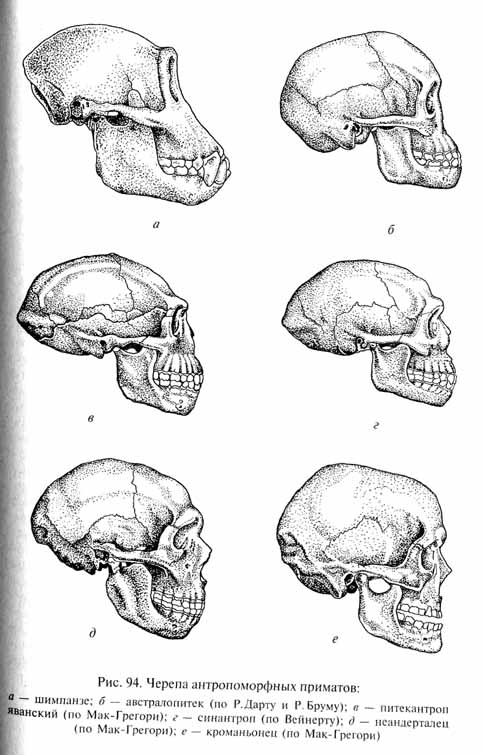
Предпосылками для развития сложных форм поведения у антропоморфов было увеличение размеров головного мозга, достигающего у шимпанзе среднего объема 405 см3, а у горилл - 498 см3, и совершенствования его структуры. Соответственно мозговая коробка получила в их черепе еще большее относительное преобладание над лицевым отделом (рис. 94).
Эволюция человекообразных приматов
Некоторые антропологи полагают, что общим предком человекообразных приматов (антропоморфов) является проконсул (Proconsul), ископаемые остатки которого были обнаружены в раннемиоценовых отложениях Восточной Африки в Кении. Проконсул был типичной мозаичной формой, в организации которой соединились некоторые признаки низших обезьян, гиббонов и шимпанзе.
Расхождение филогенетических стволов, ведущих к двум семействам антропоморфов - человекообразным обезьянам (понгидам) и людям (гоминидам), произошло, возможно, уже в миоцене (по разным оценкам от 15 до 25 млн лет назад). Из верхнемиоценовых отложений Европы известны ископаемые остатки обезьян-дриопитеков (Dryopithecus), которые по основным особенностям строения и по размерам тела напоминали современных шимпанзе. Многие антропологи рассматривают дриопитеков как возможных непосредственных предков всех высших антропоморфов, т.е. понгид и гоминид.
В неогене антропоморфные приматы достигли своего высшего расцвета. Их ископаемые остатки широко представлены в местонахождениях гиппарионовой фауны. Это говорит о связи миоплиоценовых антропоморфов с характерными для этой фауны открытыми ландшафтами, тогда как большинство приматов и особенно низшие их представители являются обитателями тропических лесов, обильных в палеогене. Вероятно, расцвет антропоморфных приматов был связан с распадением единого лесного пространства на островные леса. Можно полагать, что древнейшие человекообразные обезьяны обитали в разреженных лесах на границах лесных массивов и лесостепей. В этих пограничных местообитаниях и произошла дивергенция понгид и гоминид: первые остались лесными обитателями, предки же вторых перешли к освоению все более открытых ландшафтов. Предпосылки для этого заключались, во-первых, в переходе к наземному образу жизни; во-вторых, во все большем использовании различных орудий для зашиты и нападения и соответственно в освобождении рук от участия в передвижении; в-третьих, в совершенствовании двуногого (бипедального) передвижения. Использование орудий (грубо обработанных камней и костей животных) вместе с высокоразвитым социальным поведением позволили древним гоминидам, не имевшим ни острых клыков, ни крепких когтей, ни мошной мускулатуры (по сравнению, например, с гориллами), не только обороняться от нападений крупных хищников, но и перейти регулярному добыванию животной пищи (лишь эпизодически используемой шимпанзе).
В качестве наиболее древних и примитивных представителей эволюционного ствола гоминид многие антропологи рассматривают рамапитека (Ramapithecus) и близкого к нему сивапитека (Sivapithecus), ископаемые остатки которых были обнаружены в верхнемиоценовых отложениях Индии, с абсолютным возрастом около 12 млн лет, а также в близких по времени возникновения отложениях в Китае, Кении и Венгрии. У рамапитековых, как и у гоминид, зубные ряды левой и правой сторон сзади расходятся друг от друга, а не идут параллельно, как у большинства обезьян; клыки относительно малы, а коронки крупных коренных зубов покрыты толстым слоем эмали. Вероятно, сивапитеки и рамапитеки были уже полностью наземными всеядными обезьянами. Они широко расселились в Африке, Европе и Южной Азии в конце миоцена и существовали, вероятно, с 14 до 8 млн лет назад. Не позднее этого времени произошла дивергенция линий, ведущих к высшим человекообразным обезьянам и к гоминидам (по мнению ряда антропологов, эта дивергенция произошла даже раньше - около 15 млн лет назад, на уровне дриопитеков).
К сожалению, пока еще нет палеонтологических данных о ближайших последующих этапах филогенеза гоминид. Зато большое количество ископаемых остатков высших антропоморфов известно начиная с верхнеплиоценовых отложений, имеющих возраст от 3,5-4 млн лет и моложе. Эти остатки принадлежат различным формам африканских австралопитековых (Australopithecinae): австралопитек (Australopithecus afarensis, A. africanus), парантроп (Paranthropus robustus), плезиантроп (Plesianthropustransvaalensis), зинджантроп (Zindjanthropus boisei). Многие антропологи полагают, что все эти формы являются видами одного рода Australopithecus.
Впервые остатки австралопитеков были обнаружены в Южной Африке в 1924г. С тех пор эти антропоморфы остаются в центре оживленных дискуссий.

Австралопитеки (рис. 95), обитавшие в Южной и Восточной Африке во второй половине плиоцена и начале плейстоцена, во Многих отношениях были гораздо ближе к людям, чем любые человекообразные обезьяны. Судя по строению их таза и скелета задних конечностей, австралопитеки постоянно использовали двуногое передвижение, хотя оно было значительно менее совершенным, чем у современных людей. Пропорции их черепа (см. рис. 94) и особенности зубной системы также более сходны с человеческими, чем с обезьяньими. Средний объем мозговой коробки австралопитеков, однако, оставался близким к таковому человекообразных обезьян - в среднем 520 см3, достигая в отдельных случаях 650 см3.
Вероятно, австралопитеки достаточно часто использовали мясную пищу, охотясь на различных животных с помощью камней и тяжелых костей крупных копытных.
Размеры тела этих приматов были сравнительно невелики: по расчетам, длина их тела составляла 133-154 см при среднем весе 36-55 кг. Средняя продолжительность жизни австралопитеков оценивается А.Манном в 17-22 года. По мнению этого ученого, в жизни австралопитеков большую роль играла передача опыта от одного поколения другому, что требовало достаточно длительного периода обучения детей.
Следует ли считать австралопитеков примитивными людьми или все же лишь высокоразвитыми человекообразными обезьянами и соответственно рассматривать их в составе понгид или же гоминид? Ведь объем их мозга и ряд особенностей строения оставались гораздо более близкими к состоянию обезьян, чем людей. Дискутируется в современной антропологии и другой вопрос: были ли австралопитеки прямыми предками людей или представляли слепую боковую ветвь, развивавшуюся параллельно ветви настоящих гоминид? Некоторые ученые предлагали рассматривать австралопитеков как подсемейство в пределах семейства понгид, другие - как подсемейство в составе гоминид (эта точка зрения получила наибольшее распространение), третьи - как особое независимое семейство среди антропоморфных приматов.
Дискуссия особенно обострилась после ряда замечательных палеонтологических находок, сделанных начиная с 1959г. английским антропологом Луисом Лики, его сыном Ричардом Лики и рядом других ученых в Восточной Африке, особенно в ущелье Олдовай (северная часть Танзании) и на восточном побережье озера Рудольф. Здесь в отложениях с возрастом 1-1,8 млн лет были обнаружены ископаемые остатки высокоразвитого австралопитека - зинджантропа (A. boisei), а также другого антропоморфного примата, которому дали название "человек умелый" (Homo habilis), противопоставляя его австралопитекам как представителя настоящих людей (род Homo). Основаниями для этого были некоторые прогрессивные черты в строении скелета конечностей и несколько больший объем мозговой коробки (680-700 см3), чем у типичных австралопитеков. Вместе со скелетами зинджантропа и "человека умелого" были найдены и примитивные каменные орудия, сделанные из грубо обколотых ударами галек кварца, кварцита, лавы (так называемая "олдовайская галечная культура"), а также кости животных, часть из которых была расколота древними людьми, вероятно, для добывания костного мозга. По мнению Лики, эти орудия принадлежали не зинджантропу, а примату, названному им "человек умелый". Лики рассматривал австралопитеков как слепую боковую ветвь эволюции, представители которой сохранили более примитивное строение, чем современные им древнейшие гоминиды Homo habilis.
Однако многие антропологи не согласны с принадлежностью "человека умелого" к роду Homo, указывая на морфологическую близость этого примата к прогрессивным австралопитековым и рассматривая его как одного из их представителей. В целом нет серьезных оснований "отлучать" австралопитеков от орудий галечной культуры, тем более что примитивные орудия были обнаружены и в более древних отложениях с возрастом 2-3 млн лет.
Однако на восточном берегу озера Рудольф были обнаружены остатки австралопитеков совместно с таковыми еще более высокоразвитого, чем "человек умелый", примата с емкостью мозговой коробки 800-900 см3. В отношении этой находки уже не могло быть сомнений в ее принадлежности к роду Homo.
Судя по этим данным, типичные плейстоценовые австралопитеки действительно существовали одновременно с более прогрессивными гоминидами и поэтому не могут рассматриваться как предки последних. Тем не менее остается вполне вероятным общее происхождение всех плейстоценовых гоминид, включая австралопитеков, от каких-то позднеплиоценовых предков, которыми вполне могли быть более ранние австралопитековые, пока еще мало известные. Во всяком случае, в морфологическом отношении австралопитеки занимают промежуточное положение между высшими гоминидами и человекообразными обезьянами (понгидами) и дают представление о первом важном этапе на пути гоминизации.
Что же касается вопроса о том, рассматривать ли австралопитеков как высших человекообразных обезьян или как наиболее примитивных людей, очевидно, решить его удалось бы при наличии четких критериев, определяющих принадлежность к людям. В качестве таковых указывались: постоянное использование двуногого передвижения, выпрямленное положение тела, увеличение первого пальца руки со сложной дифференциацией его мышц, увеличение головного мозга и относительных размеров больших полушарий, некоторые особенности зубной системы. По некоторым из этих показателей австралопитеки ближе к людям, по другим - к человекообразным обезьянам, занимая в целом промежуточное положение между теми и другими. М.И.Урысон отметил отсутствие морфологических критериев, с помошью которых можно было бы однозначно определить принадлежность ископаемых скелетных остатков к людям или к высокоразвитым обезьянам, и подчеркнул, что важнейшим показателем в этом плане являются найденные вместе со скелетными остатками орудия труда. Суди по их систематическому использованию австралопитеками, последних целесообразнее рассматривать как древнейших людей. Альтернативную точку зрения аргументировал А.А.Зубов, полагающий, что критерием должна служить не сама по себе трудовая деятельность, а тот "отпечаток", который она накладывает на морфологический облик гоминид, т. е. результат приспособления к трудовой деятельности различных органов (прежде всего головного мозга и передних конечностей).
Очевидно, переход к изготовлению орудий произошел на анатомической основе, характерной для австралопитеков и еще очень близкой к таковой высших человекообразных обезьян. Вероятно, на этом этапе гоминизации главную роль сыграло не столько увеличение объема мозга, размеры которого близки у австралопитеков и понгид, сколько изменение качества его работы, связанное со значительным возрастанием числа межнейронных связей и обусловившее существенные перестройки поведения, в частности широкое использование орудийной деятельности.
Трудовая деятельность стала важнейшим фактором дальнейшей эволюции человека. Поскольку использование орудий при высокоразвитой социальности, начинавшей формироваться еще у низших антропоидов, давало человеку огромные преимущества и позволяло осваивать новые местообитания и новые природные ресурсы, естественный отбор благоприятствовал таким изменениям организации древних людей, которые способствовали совершенствованию трудовой деятельности и социального поведения. Это были в первую очередь прогрессивные изменения структуры и массы головного мозга, а также мускулатуры и скелета передних конечностей, с развитием механизмов тонкой нервной координации движений. Отбор благоприятствовал развитию у людей таких форм поведения, которые облегчали их общение в процессе трудовой деятельности, а также в организации совместных охот или защиты от нападений хищников. В связи с этим совершенствовались способы обмена информацией, в частности система звуковой сигнализации, развитие которой привело к формированию членораздельной речи, а на ее основе - второй сигнальной системы, сыгравшей столь огромную роль в развитии человеческого интеллекта и культуры.
По мнению Я.Я.Рогинского, сообщества древних людей подвергались групповому отбору, благоприятствовавшему сохранению тех коллективов, в которых преобладали более развитые в социальном отношении индивиды. Это выражалось в совершенствовании тормозных механизмов мозга, позволявших снизить взаимную агрессивность, а также в развитии свойств, способствовавших обогащению знаний на основе своего и чужого опыта.
Важнейшей чертой человеческого общества является наличие фонда социальной или культурной информации, биологически не наследуемой и передающейся от поколения к поколению посредством обучения (а на более поздних этапах развития общества кодируемой письменно) и в форме созданных предыдущими поколениями орудий труда и других материальных и культурных ценностей. Рост и развитие этого социального фонда (или фонда материальной культуры) постепенно уменьшает зависимость человеческого общества от природы. Это не могло не привести к существенным изменениям самого характера эволюционных преобразований человека. Для любой человеческой популяции фонд материальной культуры, накопленный предшествовавшими поколениями, является, в сущности, важнейшей частью среды ее обитания. Естественный отбор приспосабливал человеческие коллективы к этой их специфической среде - отбор в пользу индивидов, более способных к обучению и трудовой деятельности, и групповой отбор в пользу коллективов, в которых преобладали индивиды с более развитым социальным Поведением. Зато снижалось давление отбора на совершенствование и поддержание приспособлений, повышающих устойчивость отдельных особей к неблагоприятным факторам среды, Поскольку фонд материальной культуры, социальная организация и трудовая деятельность опосредовали влияние неблагоприятных факторов внешней среды, играя своего рода защитную роль.
С другой стороны, развитие человеческого общества все в большей степени начинает регулироваться специфическими социальными закономерностями, которые сложным образом взаимодействуют с биологическими. В полной мере эта специфика эволюции человека стала проявляться лишь на более поздних этапах его филогенеза, особенно после появления человека современного вида, Homo sapiens.
Эволюция рода Homo
Филогенетическую историю гоминид, в ходе которой постепенно формировались особенности современного человека, разделяют на ряд последовательных стадий: 1) "предшественники людей", или протоантропы; 2) архантропы; 3) палеоантропы; 4) неоантропы.
Протоантропы были представлены австралопитеками, существовавшими в позднем плиоцене и плейстоцене 5,5-1,0 млн лет назад, но, возможно, возникшими еще раньше. Австралопитеки создали костяную и галечную олдовайскую культуру. Наиболее существенные морфологические преобразования на данном этапе гоминизации происходили в скелете и мускулатуре задних конечностей. Эти изменения были связаны с переходом к постоянному двуногому передвижению. Эта стадия антропогенеза была рассмотрена нами в предыдущем разделе.
Архантропы известны по многочисленным находкам, сделанным в самых различных уголках Старого Света. Первая из этих находок была сделана еще в 1891 г. Е.Дюбуа, обнаружившим на острове Ява ископаемые остатки существа, получившего название питекантроп или "обезьяночеловек". Первоначально разные архантропы, как и австралопитеки, рассматривались как представители разных родов: питекантропы с Явы, синантропы из ряда местонахождений на территории Китая, гейдельбергский человек в Европе, атлантропы в Северной Африке и др. Ныне большинство ученых считают всех архантропов принадлежащими к одному виду Homo erectus - "человек выпрямленный", в пределах которого выделяют до 9 подвидовых форм. До недавнего времени считали, что архантропы существовали на Земле в среднем плейстоцене, со времени гюнцского и до миндельского оледенения, т.е. 700-300 тыс. лет назад, однако новые данные (например, упомянутые выше находки, сделанные Р. Лики на восточном берегу озера Рудольф) и уточнение датировок расширили время существования архантропов до огромного диапазона 2,6-0,2 млн лет назад-В течение столь длительного времени архантропы не оставались неизменными. На этой стадии антропогенеза были сделаны важные шаги по пути морфофизиологического прогресса. Так. емкость мозговой коробки увеличилась с 750-800 см3 (у древнейших архантропов, подобных лантьянскому синантропу, существовавшему около 700 тыс. лет назад, или еще более древнему питекантропу с берегов оз. Рудольф) до 1225 см3 (у синантропов из пещеры Чжоукоудянь, живших около 300 тыс. лет назад).
Однако все архантропы сохраняли целый ряд очень примитивных (для человека) признаков (см. рис. 94): очень покатый лоб, переходящий в низкий черепной свод; сильно выступающий надглазничный валик, позади которого череп был сжат поперечно (заглазничное сужение); выступающие вперед челюсти; отсутствие подбородочного выступа. Общий облик архантропов (рис. 96) был уже несомненно человеческим, хотя телосложение оставалось очень грубым, а походка была более неуклюжей, чем у современных людей.

Архантропы питались и мясной, и растительной пищей, хотя первая в их рационе, вероятно, преобладала. Вместе с остатками некоторых архантропов были обнаружены каменные и костяные орудия, более совершенные, чем олдовайские, но в целом еще очень примитивные (культура раннего палеолита). В пещере Чжо-Укоудянь (в 60 км к югу от Пекина) вместе с многочисленными Ископаемыми остатками синантропов были обнаружены каменные и костяные орудия и следы костров. Многие найденные здесь же кости животных имели следы обжига. Использование огня для обогревания пещер, служивших жилищем, и для приготовления пищи, несомненно, было важным шагом вперед, свидетельствующим о достаточно высоких познавательных и манипулятивных возможностях архантропов. Различные архантропы жили не только в пещерах, но и в лесах, и в саваннах. Вероятно, продолжительность жизни этих древних людей была небольшой; немногие из них доживали до 30-32 лет.
Следующая стадия в эволюции гоминид, палеоантропы, представлена так называемыми неандертальцами (Homo neander-thalensis), видовое название которых связано с первой находкой ископаемых остатков этих людей в долине Неандерталь вблизи Дюссельдорфа. Неандертальцы, как и архантропы, были распространены практически по всей территории Старого Света и очень разнообразны. Они появились на Земле около 300 тыс. лет назад (во время миндель-рисского межледниковья) и просуществовали до первой половины вюрмского оледенения, т. е. примерно до 35 тыс. лет назад.
Палеоантропы достигли огромного прогресса в увеличении массы головного мозга. Объем мозговой коробки мужчин-неандертальцев в среднем составлял около 1550 см3, достигая 1600 см3. Размеры головного мозга, достигнутые неандертальцами, в ходе последующей эволюции при достижении стадии неоантропов далее не увеличивались, хотя происходили перестройки структуры мозга.
Несмотря на объемистую мозговую коробку, череп неандертальцев (см. рис. 94) сохранял еще многие примитивные особенности: покатый лоб, низкий свод и затылок, массивный лицевой скелет со сплошным надглазничным валиком, подбородочный выступ был почти не выражен, сохранялись крупные зубы. Пропорции тела палеоантропов (рис. 97) были в целом близки к таковым современного человека. По сравнению с архантропами у палеоантропов усовершенствовалось строение кисти. Средний рост неандертальцев составлял 151 - 155 см.
Палеоантропами была создана культура среднего палеолита. Неандертальцы хоронили своих умерших с погребальными обрядами, что позволяет предполагать у них наличие достаточно развитого абстрактного мышления.
Стадия неоантропов соответствует человеку современного вида - Homo sapiens (человек разумный). Древнейших неоантропов традиционно называют кроманьонцами по месту первой находки их ископаемых остатков в гроте Кро-Маньон, на территории французской провинции Дордонь. Кроманьонцы уже вполне соответствовали антропологическому типу современного человека (рис. 98), отличаясь лишь незначительными особенностями (несколько менее высокий свод черепа, сильнее развитая зубная система и др.). Кроманьонцы известны начиная со времени среднего вюрмского оледенения в позднем плейстоцене около 38-40 тыс. лет назад. Однако, по некоторым данным, организация неоантропов начала формироваться еще раньше, и древнейшие неоантропы могли существовать уже 40-50 тыс. лет назад.
Средний объем черепномозговой полости у неоантропов составляет 1500 см3, т.е., как мы уже отмечали, увеличение размеров головного мозга прекратилось после достижения стадии палеоантропов. Очевидно, этот объем мозга оказался достаточным для всего последующего усложнения высшей нервной деятельности человека, вплоть до наших дней. Более того, головной мозг современного человека, объем которого не превышает такового неандертальцев, по данным физиологов, сохраняет огромные ресурсы нервных клеток, с возможностью возникновения еще большего количества нервных связей, которые остаются неиспользованными в течение всей жизни индивида (еще один пример конструктивной избыточности структур организма, см. с. 138-140).
Основные морфологические преобразования, происшедшие в процессе формирования неоантропов, выражаются в некоторых структурных изменениях головного мозга и черепа (см. рис. 94), особенно его лицевого отдела (относительное уменьшение челюстей, образование подбородочного выступа, редукция надглазничного валика и заглазничного сужения, увеличение высоты черепного свода и т.п.).
Кроманьонцы были создателями культуры позднего палеолита, характеризующейся высоким совершенством обработки камня и кости. Именно кроманьонцы были творцами пещерных рисунков, запечатлевших животных мамонтовой фауны, а также древнейших скульптурных изображений и первых музыкальных инструментов. Можно поэтому утверждать, что с неоантропами возникает искусство.
Подчеркнем еще раз, что каждая из рассмотренных нами стадий эволюции человека включала большое количество вариаций - как в пространстве (в разных регионах), так и во времени. Характерные особенности следующей стадии не возникали внезапно и все сразу, но постепенно развивались у разных популяций, так сказать, "в недрах&
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





