|
|
Открытие мобильных элементов и непостоянства геномаЯвление нестабильности наследственного фактора первым обнаружил де Фриз (1901), когда описал мозаичность окраски цветка львиного зева. Нестабильность стала одним из подтверждений гипотезы о мутационных периодах. Разумеется, о познании ее генетической природы тогда речи идти не могло. Приоритет в открытии мобильных генетических элементов (МГЭ) и нестабильности генома принадлежит по праву американскому генетику, специалисту по цитогенетике кукурузы Барбаре МакКлинток. Она тоже обратила внимание на мозаичность окраски зерен у этой культуры. В серии работ 1951—1965 гг. МакКлинток выдвинула гипотезу о существовании особого класса контролирующих подвижных элементов, способных перемещаться по геному, встраиваться в разные локусы (равно как и удаляться из них) и таким путем регулировать темп мутирования гена и его мутационное состояние. МакКлинток показала, что мутации при этом могут возникать с частотой, в сотни и тысячи раз превышающей обычную, и что1 они носят упорядоченный характер. МакКлинток совмещала в работе генетический и цитогенетиче-ский подходы и в наблюдениях под микроскопом убеждалась в правильности ожиданий, вытекавших из генетического анализа.
Из наблюдений МакКлинток следовало, что мутационное событие может быть связано не с изменением самого гена, а с неким подвижным контролирующим его элементом, способным вызывать разрывы в местах внедрения и встраиваться в разные локусы. Ею также было замечено, что разрывы хромосом обычно происходят в определенных сайтах, хотя их топография могла и изменяться. Хотя сообщения о результатах первых исследований МакКлинток были напечатаны в самь|Х авТ°Ритетных американских научных изданиях (McClintock, 1950, 1951), а она сама была избрана членом Американской академии наук и пользовалась репутацией признанного специалиста, ее открытия казались какой-то абсурдной экзотикой, совершенно чуждой привычному представлению моргановской генетики о постоянной прописке генов. Их постигла та же судьба, что и открытие Менделя в XIX в. Они были восприняты и оценены лишь через 25 лет, когда для этого созрели необходимые условия. Тем временем в разных областях генетики множились данные, свидетельствовавшие о реальности существования подвижных генетических элементов, призванных осуществлять регулирующую функцию в отношении «постоянных» генов. Лишь к концу 70-х годов они перестали выглядеть разрозненными фактами и соединились в единой концепции. В 60-е годы у бактерий был обнаружен класс инсерционных мутаций, вызванных внедрением чужеродных сегментов ДНК, а затем были выделены и сами инсерционные элементы. Удалось получить доказательства инсерционной природы нестабильных мутаций и у лабораторных линий дрозофилы (Green, 1967, 1969). Американский генетик М. Грин одним их первых показал, что в природных популяциях дрозофил существуют генетические элементы, которые индуцируют инсерционный мутагенез во множестве ло-кусов (см.: Golubovsky, Ivanov, Green, 1977). Исследования спонтанного мутационного процесса в природных популяциях дрозофил имели для доказательства существования МГЭ особое значение. Еще в 30-х годах Р.Л. Берг наблюдала сначала резкую вспышку мутабельности гена «желтое тело*, а затем, в конце 40-х годов, -— восстановление нормального уровня мутирования. В 1968 г. то же наблюдалось в отношении мутации «abnormal abdomen», а в 1973 г. — для гена singed, сцепленно- го с полом (Berg, 1974, 1982; Голубовский и др., 1974). Аллели этого последнего гена, выделенные из различных популяций в период вспышки, оказались нестабильными: они, в частности, мутировали к норме с необычно высокой частотой. Впервые в природных условиях наблюдалась вспышка целой серии нестабильных аллелей. Генетический анализ, проведенный Голубовским с соавт. (Golubovsky, Ivanov, Green, 1977; Golubovsky, 1980), однозначно показал, что в данном случае имел место инсерционный мутагенез. Голубовский (2000) отмечает, что поведение нестабильных природных аллелей дрозофилы оказалось весьма сходным с поведением нестабильных генов у кукурузы. Переломным в борьбе за признание МГЭ можно считать 1978 г. Об открытии подобных элементов у дрозофилы и млекопитающих сообщила лаборатория Г.П. Георгиева в Институте молекулярной биологии бывшей АН СССР и на дрозофиле — группа исследователей в США во главе с Г. Хогнесом (Ilyin et al., 1978; Finnegan et al., 1978). О своем приоритете напомнила МакКлинток (McClintock, 1978). Лед тронулся. Прошло еще немного времени, и признание МГЭ defacto, подтвердившее соображения о непостоянстве генома и горизонтальном переносе генов (Хесин, Кордюм), было воспринято как настоящая революция в молекулярной генетике. Почему же пионерские работы МакКлинток столь долгое время оставались на периферии генетики? Голубовский (2000), уделивший этому вопросу большое внимание в своей книге, дает на него ясный ответ: идеи МакКлинток были для концептуального поля классической генетики и СТЭ инородным телом. Ее выводы противоречили основным постулатам хромосомной теории наследственности, таким, как постоянство положения гена в хромосоме, случайность мутаций, их низкая частота и непредсказуемость. Из работ МакКлинток, напротив, следовало, что активность генов находится под контролем регуляторных факторов, что их мутации могут возникать с большой частотой и упорядоченно, что число самих МГЭ широко варьирует. Хочется воспроизвести фрагмент из последней печатной работы МакКлинток, в котором она дистанцируется от СТЭ и солидаризуется с Гольдшмидтом. «Не вызывает сомнения, — писала она, — что гены некоторых, если не всех организмов лабильны и что резкие их изменения могут происходить с большой частотой. Эти изменения могут вести к реорганизации генома и к изменениям в регуляции активности и времени выражения гена. По- скольку способы реорганизации генома за счет мобильных элементов разнообразны, их активация, за которой следует стабилизация, может дать начало новым видам и родам» (McClintock, 1984; цит. по: Голубовский, 2000. С. 215). Изучение МГЭ, или, как их еще стали называть, транспозиционных элементов (TEs), продвигается быстрыми темпами. Эти элементы представляют собой частицы ФК генома, представленные эгоистической ДНК, число и топография которых уникальны для каждой особи. Они действуют как самопроизвольные мутанты и производят в основном рецессивные мутации. После всеобщего признания МГЭ выяснились их широкое распространение в природе и способность к сальтационным эволюционно-генетическим перевоплощениям в ряду: простые инсерционные последовательности — транспозоны — плазмиды — вирусы. На сегодняшний день у дрозофилы известно свыше 30 семейств МГЭ. Их доля в геноме DrosophiJa melanogaster достигает около 15 % {Голубовский, 2000). Из числа спонтанных мутаций у этого вида 70 % связаны с инсерциями, которые дают разные результаты. При инвазиях МГЭ наблюдаются как вспышки мутаций в популяциях, так и массовые определенные изменения. Допускается, что упорядоченные перестройки генома с участием МГЭ могут программироваться и запускаться на определенных этапах онтогенеза. Таким путем преадаптационно создается, например, аллельное разнообразие генов иммунной системы. Активизация некоторых семейств подвижных элементов способна вызывать у насекомых и комплекс разнородных явлений, которые известны под названием гибридного дисгенеза (впервые описан Kidwell et af., 1977), ныне пристально изучаемого во многих лабораториях мира. Примером могут служить исследования лаборатории М.Б. Евгеньева в Институте молекулярной биологии, работающей с древней группой Drosophila virilis. Группе Евгеньева удалось выяснить, что за гибридный дисгенез у них ответственен элемент Penelope. Элемент обладает чрезвычайно сложной и крайне изменчивой организацией, способен передаваться как вертикально (от поколению к поколению), так и горизонтально и активировать другие транспозиционные элементы. Было также установлено, что на протяжении существования Dr. virilis Penelope вторгался в его популяции несколько раз. В мировой литературе имеются также свидетельства, что ряд транспозиционных элементов проник в популяции Drosophila melanogaster в XX в. посредством горизонтального переноса от других видов {Kidwell, 1983; Bucheton et al., 1992; Simmons,1992). Сказанного здесь, а также в главе о чужеродных генах вполне достаточно, чтобы убедиться в совершенно исключительных и безграничных возможностях мобильных элементов, которые делают их в наших глазах повсеместными и всепроникающими генетическими агентами, генераторами всевозможных и поистине волшебных изменений. Универсальное значение мобильных элементов для преобразования структуры генома и расширения эволюционных потенций их обладателей делает понятным, почему современную неклассическую генетику в обиходе называют также «подвижной». Стресс и генетический поиск
Еще в первой половине XX в. физиологи столкнулись с неспецифической адаптационной реакцией животных, развивавшейся в ответ на неблагоприятные воздействия внешней среды (холод, голод, травмы) или негативные психические состояния (страх, тревога и пр.). Реакции затрагивали сферу нейроэндокринной регуляции и были связаны с изменением режима функционирования желез внутренней секреции. Канадский патолог Ганс Селье, впервые описавший этот феномен под названием стресса (Selye, I936), на протяжении многих десятилетий разрабатывал одноименную физиологическую концепцию (Selye; 1950,1956; Селье, 1960, 1972, 1979), и она обросла огромной литературой.
Селье определил стресс как «совокупность всех неспецифических изменений, возникающих под влиянием любых сильных воздействий и сопровождающихся перестройкой защитных систем организма» (Селье, 1972. С. 116). Впоследствии он дал ему более общую дефиницию: «Стресс есть неспецифический ответ организма на любое предъявленное ему требование» (Селье, 1979. С. 27). Главной причиной возникновения стресса, по Селье, служит отклонение любого жизненно значимого параметра внутренней или внешней среды организма от оптималього уровня, нарушающее его гомеостаз. Последователи Селье распространили понятие стресса на все живое и сочли полезным и необходимым изучать ответные реакции организма на всех уровнях его организации — вплоть до молекулярного. Стресс — это испытание. При экстремальных значениях и большой продолжительности действия стрессорных факторов организм погибает, а популяция сокращает свою численность практически до нуля. При более умеренных параметрах воздействия стресс становится механизмом выживания, ибо стимулирует поиск полезных защитных реакций и нужных форм поведения. Как показали полевые и лабораторные исследования, целесообразные изменения в поведении в состоянии компенсировать рост энергетических затрат организма при стрессе от абиотических воздействий. Замечено также, что способностью к поддержанию более эффективного энергетического баланса в большей мере обладают гетерозиготы (Parsons, 1996; Парсонс, 2000). Хотя стресс не является приспособлением к определенному фактору среды, не вызывает сомнений, что этот синдром — одно из проявлений адаптивной нормы, возникшее в ходе эволюции и детерминируемое системой онтогенетического развития. Наряду с физиологическим существует стресс геномный. Как показывает название, речь идет в этом случае о реакции на те же стрессорные факторы генома организма, который может испытывать быструю и существенную реорганизацию. Побуждение исследователей к изучению этой формы стресса опять-таки связано с работами МакКлинток. Последняя не только открыла МГЭ, но и убедительно показала, что эти элементы составляют часть системы, с помощью которой живая клетка способна целенаправленно перестраивать свой геном в ответ на стресс (McClintock, 1978, 1984). В нобелевской лекции (1984) МакКлинток указала также, что эта перестройка может служить основой образования новых видов. Между физиологическим и геномным стрессами имеется не только смысловое сходство. Скорее всего обе эти формы стресса составляют единый адаптационный механизм, направленный на защиту организма от тех факторов, с которыми его вид никогда не сталкивался за свою эволюционную историю либо при которых не происходило адекватной адаптивной реакции. Обе реакции неспецифичны, но при этом строго упорядоченны. По мнению Ю.И. Аршавского2, организм сам ищет и находит нужное
изменение своей физиологии, которому затем подыскивается подходящее генетическое основание. Главной чертой геномного стресса можно считать усиление наследственной изменчивости, отмечаемое многими авторами (Parsons, 1988, 1993, 1996; Carson, 1990; Чайковский, 1998, 2001). При этом возрастает частота мутаций и рекомбинаций, увеличивается вариабельность онтогенеза и многих фенотипических признаков. Но мутируют очень немногие гены и в весьма ограниченном числе направлений. Для обозначения генетических изменений, происходящих при стрессе, Ю.В. Чайковским был предложен удачный термин — генетический поиск. Под этим названием Чайковский понимает «тот исключительный режим работы генетической системы, в котором изготавливаются новые генетические тексты (в обычном режиме генетическая система лишь копирует и комбинирует прежние тексты)» (Чайковский, 1976. С. 156-157). Он особо отмечает при этом, что новые тексты ДНК должны создаваться и перестраиваться по каким-то достаточно жестким законам. Клетка с ее системой наследственной памяти способна ответить на вызов среды активным и упорядоченным генетическим поиском, а не пассивно «ждать» случайного возникновения адаптивной мутации. Голу-бовский (1999) уверен, что появление новых генов должно сопровождаться изменениями в количественном составе и внутригенной топографии разных факультативных элементов и образованием новых наследуемых эпигенетических систем регуляции. В наши дни все чаще полагают, что главным поставщиком изменчивости являются скорее не мутации отдельных генов, а рекомбинации целых блоков генетического материала. Селекция их лучших вариантов осуществляется на клеточном уровне. Появились веские свидетельства, что важная роль в дестабилизации и перестройке генома принадлежит МГЭ (Junakovic et al., 1986; Anaya, Roncero, 1996; Васильева и др., 1997). Перемещения МГЭ неслучайны, а места их встраивания в хромосомы специализированны. Если угодно,генетический поиск есть одно из выражений постулата о наследовании приобретенных признаков. Желая разобраться, как работает подобный механизм, ищущий ум более, чем где бы то ни было еще, сталкивается с ощущением недостаточности, а то и непригодности используемых механизмов познания. В исследованиях на растениях показано (Hollick et al., 1997; Кал-лис, 2000), что быстрые реорганизации генома связаны с количественными изменениями повторяющейся ДНК, с ее метилированием, с инсерциями МГЭ, амплификацией или делецией генов. 41S Однако при определенных успехах в познании механизмов генетического поиска мы еще мало что о них знаем (Маркель, 2000). В качестве одного из возможных сценариев генетического поиска приведем отрывок из работы Голубовского (1985а), хорошо отражающий объяснительные возможности новой генетики. Стресс, при котором, как уже говорилось, активность генов в ФК повышается, может сопровождаться их магнификацией и увеличением вероятности включения их дополнительных копий в ОК. Магнификация совершенно очевидна в случае таких жизненно важных генов, как рибосомные и гистонные. Если вызвать генетический стресс, уменьшив с помощью делеции дозу рибосомных генов на 50 %, то в соматических клетках их доза восполняется за одно поколение за счет амплификации и образования внехро-мосомных копий. Если число рибосомных генов сократить еще на 20 %, то число генов восстановится до нормы не только в соматических, но и в половых клетках, В этих последних процесс протекает ступенчато, за три-пять поколений. Вначале магнифи-цированные копии генов в половых клетках наследуются нестабильно, и, если стрессовое давление снимается, происходит быстрая реверсия к норме. Система как бы проверяет, насколько серьезны и устойчивы неблагоприятные воздействия. Но, если они действуют долго (более пяти—семи поколений), происходит стабильное включение магнифицированных копий в состав ОК. Описанный процесс носит направленный, определенный и постепенный характер, он захватывает сразу большое число особей, чем принципиально отличается от мутаций. Это тот тип наследования, который наблюдается в случае длительных модификаций. В Советском Союзе большие формообразующие и эволюционные возможности стресса продемонстрировали многолетние работы Д.К. Беляева (1970, 1977, 1979а, б) по селекции серебристо-черных лисиц на доместикационный тип поведения. Беляев и руководимый им коллектив в Институте цитологии и генетики СО АН СССР установили, что стресс в кратчайшие сроки резко повышал наследственную изменчивость и дестабилизировал системы онтогенеза. При этом наблюдалась мобилизация резерва доселе не проявлявшихся мутаций, в стресс вовлекался весь генетический аппарат, контролирующий нейроэндокринные механизмы процессов развития, со всеми вытекающими отсюда последствиями. Важнейшим из них оказалось резкое увеличение темпов формообразовательных процессов. Результаты отбора на стрессо-устойчивость (Беляев назвал его дестабилизирующим) убедили Бе- ляева в том, что «стресс служит важнейшим модусом эволюции, ее фактором» (Беляев, 1979а. С. 45). К достижениям новой генетики можно отнести установление принципа блочной организации хромосом эукариот. Начало было положено еше К. Дарлингтоном и К. Мазером. Швейцарский генетик В. Геринг открыл в составе регуляторных, гомеозисных генов общий сегмент — гамеобокс, способный регулировать активность целой группы структурных генов. К настоящему времени установлено, что гены вообще чаще всего объединены в кластеры (а соответствующие белки — в домены), отделенные друг от друга промежутками. Таким способом, в частности, организовано семейство глобиновых генов человека. В хромосоме I i в районе протяженностью 60 тыс. п. н. расположено 7 несколько отличающихся друг от друга копий бета-глоби-новых генов (5 из них функционально активны на разных стадиях онтогенеза). В хромосоме 16 расположено другое семейство из пяти альфа-глобиновых генов. Блочная организация генома обеспечивает развивающемуся организму функциональные преимущества. Разные генные блоки активны в разных тканях или на разных этапах онтогенеза, и благодаря такому объединению генов целые их семейства могут быть сразу включены или выключены. Этот же технологический принцип используется, очевидно, и в эволюции. По представлениям Корочкина (1984, 1999), он облегчает смену программ индивидуального развития, а по Кордю-му (1982) — поступление и элиминацию экзогенной генетической информации. В общем виде идею эволюции как преобразование комбинации блоков на примере совершенствования пищеварительной системы развил петербургский физиолог A.M. Уголев (1985). Уголев видит и находит функциональные блоки на всех уровнях организации — биосферном, ценотическом, популяционном, ор-ганизменном, клеточном, субклеточном — и считает, что наиболее быстрый и эффективный способ эволюции состоял в смене их комбинаций (Уголев, 1994). Подобные же идеи разделяют Мейен и Чайковский (1990, 2000). Мы рассмотрели далеко не все открытия и концепции, которые составили основу современной «подвижной» генетики, ограничившись теми, что наиболее существенны для понимания видообразования и эволюции. В заключение главы приведем таблицу 3, в которой в тезисной форме сопоставлены представления классической и современной генетики о наследственной изменчивости. Таблица 3. Сопоставление представлений о наследственной изменчивости в классической и современной генетике (из: Голубовский, 2000)
Глава 17 ГЕНЕТИКА И ЭПИГЕНЕТИКА ВИДООБРАЗОВАНИЯ И МАКРОЭВОЛЮЦИИ Ушедший XX век подвел науку об эволюции к парадоксальной ситуации. Генетика с ее специализированными материальными носителями наследственности, к открытию которых упорно стремились многие поколения исследователей, те самые молекулы ДНК, которые еще недавно царствовали в умах биологов-эволюционистов как азбука любых перемен в живом, вдруг оказались малозначимыми для теории эволюции! У эволюции свои законы, и генетическая система у них всего лишь в подчинении. Зато эволюция, если говорить образно, шагает под руку с эпиге-нетикой и даже позволяет ей время от времени себя вести. Между тем эпигенетика пользуется языком генетики и считает для себя необходимым изучать все, что происходит в геноме. Причина столь разного отношения эволюции и эпигенетики к генетике ясна: эволюция вполне «сознает», что генетический аппарат — ее порождение, но никак не причина. Тогда невольно возникает вопрос: зачем ученым-эволюционистам интересоваться генетическими изменениями, если они не первичны? Вероятно, данный парадокс способен вызвать то же замешательство, какое может испытывать здравомыслящий гражданин, впервые увидевший уличный щит с рекламой сигарет, а под ней неброское предупреждение Минздрава о том, что курение опасно для здоровья. В данном случае, как нам представляется, разумно опираться на следующее соображение. Претерпевая те или иные изменения и осуществляя генетический поиск, генетический аппарат создает базу для надежного воспроизведения тех удачных поведенческих и физиологических реакций, которые осуществил организм в ответ на сдвиги во внутренней или внешней среде. Перед нашим взором прошло достаточно примеров крушения разных догм и «возвращения к истокам». Не будем удивляться и упомянутому парадоксу и воспримем его с надеждой, что в будущем наука сможет его удовлетворительно разрешить. Смиримся также с фактом, что при всех успехах генетики и селекции мы не можем похвастаться созданием сколько-нибудь значительного числа полноценных биологических видов1. С точки зрения наших
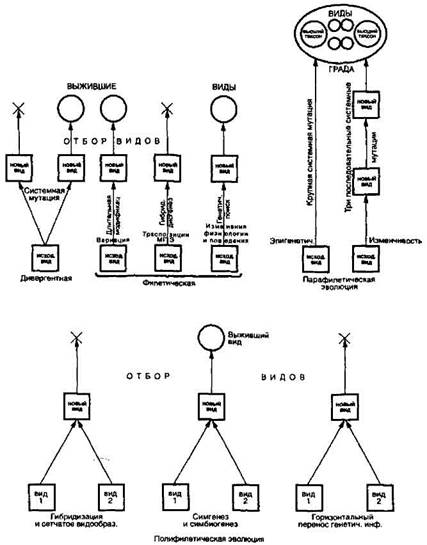
познавательных возможностей это вызывает разочарование, но зато служит дополнительным стимулом к созданию истинной и, значит, действенной теории видообразования. Обратимся теперь к тому, что известно из области эпигенетических и генетических событий, сопутствующих образованию видов и более высоких таксонов. Только будем помнить, что генетические изменения не диктуют организму, что ему делать, а всего лишь фиксируют в своей генетической памяти его правильные эволюционные шаги. За последние 30 лет здесь произошла еще более радикальная революция, чем в молекулярной генетике. Прежние представления о способах видообразования, основанные на модели СТЭ, оказались совершенно несовместимыми ни с теорией прерывистого равновесия, ни с эпигенетикой, ни с новой генетикой. На смену им пришло множество альтернативных гипотез, часть из которых успели обрести статус достаточно прочно обоснованных теорий. Все эти годы объем непредвзятых исследований по проблеме видообразования продолжал нарастать, но все попытки выяснить, какие именно генетические изменения (и в каком количестве) связаны с видообразованием, до сих пор не привели к какой-либо единой схеме. И это вполне объяснимо, ибо в связи с различиями в строении генома, в детерминации и течении процессов индивидуального развития, в способах размножения и т. п. у разных групп организмов и не может быть единого механизма видообразования. Не приходится удивляться, таким образом, что органический мир демонстрирует нам множественность путей и способов образования новых видов, с некоторыми из которых мы уже знакомы. Разнообразие, однако, не исключает принципиальной общности. Главные способы видообразования, которые мы рассмотрим ниже, отвечают следующим характеристикам. Новые вилы образуются только сальтационно в течение одно-го-двух или ограниченного числа поколений в результате установления репродуктивной изоляции. При такой скорости видообра-зовательного акта захват популяции новым видовым комплексом признаков путем вытеснения старого на основе генетико-популя-ционных процессов невозможен, так как такие процессы протекают слишком медленно и обычно не имеют завершения. Поскольку популяционные процессы к видообразованию не ведут, естественный отбор оказывается без объекта своего действия. К тому же возможные эффекты отбора резко подавлялись бы системой активных реакций организма. Вид рождается сразу в большом числе особей, реже — в нескольких. Изменения, осуществляемые
Рис. 27. Обобщенная схема, иллюстрирующая множественность путей видообразования (ориг.) организмом, часто носят адаптивный и направленный характер и оказываются в разных таксонах параллельными, что свидетельствует о закономерности их возникновения и справедливости номо-генетической концепции. Если изменения носят иной характер, то равновесие со средой устанавливается преадаптационным путем. Особо подчеркнем, что приведенная характеристика видообразования, каковой она видится в свете новой генетики, всецело соответствует теории прерывистого равновесия (см. гл. 15). В синтетической теории считается, что видообразование — сложный и длительный процесс, не сводимый к изменчивости. Последняя служит всего лишь «сырым материалом» для отбора, который выступает единоличным творцом новых форм. Все попытки при объяснении видообразования ограничиться одной изменчивостью, проигнорировав отбор, адептами СТЭ всегда резко осуждались. Сейчас нам совершенно ясно, почему механизмы формообразования нельзя, с позиций СТЭ, свести к изменчивости: механизм естественного отбора и целесообразные действия организма — это разные и несовместимые способы эволюции. При первом способе изменчивость безлика (неопределенна), при втором — выражение целесообразных реакций. В новых теориях и гипотезах видообразования, как правило, обходятся без отбора и действительно сводят этот процесс к наследственной изменчивости. При этом авторы теории вовсе не считают изменчивость простым процессом. Мы убедились, что ей могут соответствовать сложные генетические и эпигенетические процессы и, как считают некоторые специалисты, и она может быть результатом своеобразных внутри геномных популяционных взаимодействий. Короче, нет ничего научно порочного или методологически предосудительного, чтобы считать носителей вариационной или динамической изменчивости новыми видами, если они этого заслуживают. Системные мутации Наиболее фундаментальное значение для теории видообразования имело, с нашей точки зрения, открытие отечественными исследователями Ю.П.Алтуховым и Ю.Г. Рычковым (1972) моно-морфной части генома, о чем уже говорилось в общеэволюционном плане в гл. 3 и 15. Обнаружение мономорфных белков — это признание существования видовых (и более высоких категорий) признаков и еще одно свидетельство живучести идей Ламарка, который первым выделил конституционные признаки организмов по их особой значимости для эволюции. Это также лишнее подтверждение правильности применения к видообразованию типологического подхода. В гл. 15 говорилось, что реорганизация вилоспецифической части генома, с точки зрения Алтухова, выражается в системной мутации и такую мутацию можно считать центральным генетическим событием. Не вызывает сомнения, что само понятие системной мутации стало одной из ведущих эволюиионно-генетических идей последних десятилетий. Гольдшмидт, Вандель, Дальк и Уоддингтон, открывшие системные мутации чисто умозрительным путем, были твердо убеждены в их реальности, но не располагали доказательствами. Впервые мутации такого рода обнаружил и описал у малярийных комаров томский генетик В.Н. Стегний (1979). Они оказались в основном аналогичны системным мутациям, прокламированным Гольдшмидтом, и также касались архитектоники хромосомного аппарата, почему Стегний оставил за ними прежнее название, хотя, по сути, описанные им мутации ближе онтомутациям Далька. Новый тип системных мутаций, о котором идет речь, вписывается в концепцию мономорфизма Алтухова—Рычкова. Это базовый видоспецифический признак, не обнаруживающий внутривидового (популяционного) полиморфизма и выявляемый только в геномах генеративной ткани (в трофоцитах). При данном типе мутаций не меняется ни число хромосом, ни их линейная структура, ни генный состав. Системные мутации возникают в результате реорганизации хромосом в интерфазном ядре путем изменения хромосомно-мембранных отношений. На хромосомах и на ядерной мембране появляются (или исчезают) тяжи р-гетерохроматина, которые прикрепляют их друг к другу. Мутации связаны также с динамичным изменением хромоцентра — от локального до диффузного его размещения на периферии ядра. При всех преобразованиях этого рода про-исходит устойчивое и необратимое изменение функционирования видового генома и устанавливается межвидовой хиатус (Стегний, 1993, 1996). Автор специально подчеркивает, что ни в одном из наблюдений ни на комарах, ни впоследствии на дрозофиле (Стегний, Вассерлауф, 1991а, б) не было зарегистрировано внутривидового полиморфизма (гетерозиготности) по архитектонике генома, что однозначно свидетельствует о его исключительно сальтационном преобразовании. Градуальная реорганизация описанных хромосомно-мембранных отношений принципиально невозможна. Обнаруженные системные мутации выявили зависимость потенциальной способности видообразования от внутренней организации ядерно-хромосомного аппарата и, в частности, степени , 423 «жесткости» трехмерной структуры ядра. По этому показателю, а также по распределению гетерохроматина и инверсионного полиморфизма видовые геномы могут быть отнесены к двум категориям. Лабильный, т.е. способный к образованию дочерних видов, геном обладает следующими чертами: наличием локализованного в центральном участке гетерохроматина (1); облигатным хромосомным мономорфизмом (2); ограниченным (лучше минимальным) числом хромосомно-мембранных связей в интерфазных ядрах клеток генеративной ткани (3). Консервативный, т.е. препятствующий видообразованию, геном имеет противоположные характеристики. Гетерохроматин у него диспергирован (1), часто имеет место инверсионный полиморфизм (2), хромосомно-мембранные связи многочисленны. Необратимость видообразова-тельных событий как раз и связана с невозможностью обратного перехода гетерохроматиновых блоков из диспергированного состояния в локализованное, а у хромосом — из состояния прикреплен-ности к мембране в свободное (Стегний, 1993. С. 87-88). Предполагается, что внутренними причинами, предрасполагающими геном к системной мутации, могут быть его лабильная структурная организация и переход организмов к инбридному размножению. Решающими внешними причинами выступают экстремальные абиотические условия, и прежде всего температура. Они могут возникать как на периферии ареала, так и в его центре. В этих условиях резко снижается численность вида, а это по причинно-следственной цепочке ведет к близкородственному размножению, активизации МГЭ, стимуляции разных типов мутагенеза. Все это дестабилизирует геном. Что происходит далее? Инбридинг приводит к тому, что системные мутации появляются кластером. Последний, по наблюдениям Стегния, в течение 1-2 поколений проходит стадию гете-розиготности и фиксируется в гомозиготном состоянии. Гетерозиготы не в состоянии надолго задержаться в популяции ввиду структурной дезинтеграции их хромосом, как это отмечается у межвидовых гибридов. Гомозиготы по системной мутации, более способные к выживанию, быстро размножаются и формируют популяцию нового вида, которая занимает освобождающуюся адаптивную зону. Обратим внимание, что при сальтационном возникновении нового вида через преобразование мономорфной части генома (в том числе путем системных мутаций) отпадает необходимость в пространственной изоляции популяций и исчезает принципиальное различие между аллопатическим и симпатрическим способами видообразования. Выделение этих способов утрачивает смысл. Видообразование путем системных мутаций, кроме того, снимает проблему аллельных и хромосомных мутаций, возникающих с высокой частотой и сопряженно с системными мутациями. Поскольку последние не могут существовать в гетерозиготном состоянии более 1—2 поколений, все точковые мутации и хромосомные перестройки типа инверсий вынужденно переводятся в облигат-но-гомозиготное состояние. Таким образом, популяция дочернего вида в момент своего рождения будет мономорфной по всем генным локусам и хромосомам. Развертывание адаптивного полиморфизма (если он свойственен родоначальному виду) станет содержанием последующего стабилизационного периода. Сказанное выше позволяет по-иному взглянуть на старые эволюционные модели квантовой эволюции Симпсона и генетической революции Майра, происходящие в малых периферических изолятах и при реализации «принципа основателя». Указанные авторы приписывали эффект этих моделей изменениям внутренней генетической среды, которые влекут изменения селективной ценности многих генов. Кроме того, Майр связывал видообразование путем генетической революции с аллопатрической моделью, при которой географическая изоляция предшествует репродуктивной изоляции. В свете новых открытий в генетике нетрудно понять, что изменилось в генетической интерпретации данных модусов. Периферические изоляты действительно создают благоприятные условия для видообразования, но этот процесс сопряжен не столько с генными мутациями, сколь многочисленными они бы ни были, сколько с появлением системных мутаций, при которых репродуктивная изоляция первична, гомози готизация очень быстра, а носителей мутаций много. В трактовке системных мутаций Стегнием есть некоторые расхождения с Гольдшмидтом. Стегний считает, что системные мутации (и, соответственно, новые виды) могут вообще не иметь фенотипического (морфологического) выражения или обладать минимальными морфологическими отличиями подобно видам-двойникам. Зато возникающие на их основе новые виды всегда отличаются от исходного родительского вида физиологически. Системные мутации возникают не в виде единичной особи, как полагал Гольдшмидт, а целым пакетом (кластером), что намного увеличивает их шансы закрепления. Наконец, Стегний, в отличие от Гольдшмидта и большинства своих единомышленников, подчеркивает, что придает большое значение естественному отбору на всех стадиях видообразования, хотя в своих главных публикациях предметно этого вопроса не касается. Для окончательного упрочения концепции видообразования с участием системных мутаций было бы неплохо получить их экспериментальным путем. К сожалению, предпринимавшиеся в этом направлении попытки пока успехом не увенчались. Известную аналогию системным мутациям представляет уже известный нам гибридный дисгенез, вызываемый транспозиционными элементами. Он состоит в появлении аномального потомства от скрещивания самок Drosophila melanogaster, длительное время разводимых в лаборатории, с самцами диких популяций того же вида. В таком потомстве из-за нарушений в развитии зародышевых клеток обоих полов наблюдаются высокая стерильность, повышенная генная и   ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|


 0 Устное сообщение Ю.И. Аршавского. 414
0 Устное сообщение Ю.И. Аршавского. 414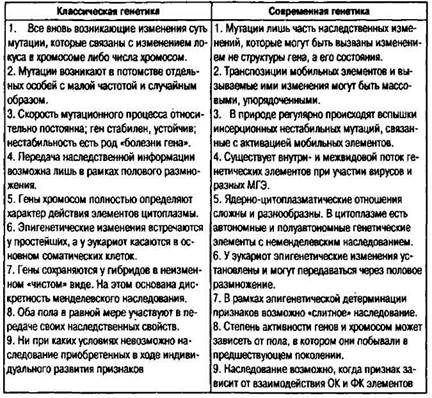
 1 Несмотря на все возможности генетической инженерии человек располагает реальной возможностью создавать виды только с помошью полиплоидии.
1 Несмотря на все возможности генетической инженерии человек располагает реальной возможностью создавать виды только с помошью полиплоидии.