|
|
Тема1. Психофізіологія як наука.Стр 1 из 6Следующая ⇒ Психофізіологія
Тема1. Психофізіологія як наука. Предмет, задачі та методи психофізіології. Зв’язок психофізіології з істотними та гуманітарними науками. Сучасні напрямки досліджень з психофізіології.
КОНСПЕКТ
Тема 2. Основні поняття та принципи роботи ЦНС та ВНД. Основні положення теорії рефлекторної діяльності. Функціональна організація мозку. Основні стуктури головного мозку. Загальні характеристики будови та роботи сенсорних систем. Теорія системної динамічної локалізації А.Р.Лурія. Конспект Структуры головного мозга человека ЦНС в значительной степени обособлена от окружающих тканей и органов тела и имеет собственную, специфическую внутреннюю среду. Головной и спинной мозг окружены системой защитных структур, в том числе тремя мозговыми оболочками. Наружная (durа таter), плотная и прочная, образует как бы герметизирующий чехол; внутренняя (рiа тater), мяг- кая, сосудистая, содержит кровеносные сосуды, питающие мозг; и промежуточная (агасНпогс1еа таСег), в виде ажурной паутинки, разделяет их. Межоболочечное пространство и внутренние полости мозга (желудочки) заполнены спинномозго- вой жидкостью (Ндиог), специфику и константность состава которой определяет сложный морфофункциональный комплекс клеточных структур, получивший общее название — «гематоэнцефалический барьер».
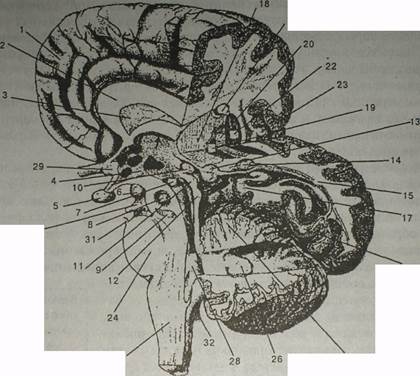
Рис. Головной мозг (правое полушарие, частично удалены теменная, височная и затылочная области) I — медиальная поверхность лобной области правого полушария; 2 — мозолистое тело (согрив саНовит); 3 — прозрачная перегородка (вер(ит реНис'кЗит); 4 — ядра гипоталамуса (гшс/е/ Луро-гЛа/ат/); 5 — гипофиз (ЬурорЫв!в); 6 — мамилярное тело (согрив татШаге); 7 — субталамическое ядро (пис!еив виЬ-(На1ат!сив); 8 — красное ядро (пис!еив гиЬег) (проекция); 9 — черная субстанция (виЬ$(ап11а п/§га) (проекция); 10 — шишковидная железа (согрив р/пеа/е); I1 — верхние бугорки четверохолмия (соШсиН вирег!ог 1ес11 тевепсерНаН); 12 — нижние бугорки четверохолмия (соШсиН т(епог 1ес(1 тевепсерНаИ); 13 —медиальное коленчатое тело (МКТ) (согрив§еп!си1а1ит тесИа!е); 14 — латеральное коленчатое тело (АКТ) (согрив §еп1си1а1ит 1а1ега1е); 15 — нервные волокна, идушие от АКТ в первичную зрительную кору; 16 — шпорная извилина (ви!сив са/саг1пив); 17 — гиппокампальная извилина (%1ги5 ЫрросатраНв); 18 — таламус ((На1атив); 19 — внутренняя часть бледного шара (@1оЬи$ ра!Н-с!ив); 20 — наружная часть бледного шара; 21 — хвостатое ядро (пис!еив саи- с/аа/5); 22 — скорлупа (ри(атеп); 23 — островок (тяи/а); 24 — мост (рот); 25 — мозжечок (кора) (сегеЬеНит); 26 — зубчатое ядро мозжечка (пис!еив с!еп(а1ив); 27— продолговатый мозг (тес!иПл оЫоп%з1а); 28 — четвертый желудочек (уеп(пси!ив ^иа^^ив)•, 29 — зрительный нерв (пегуив орИсив); 30 — глазодвигательный нерв (пегуив оси1опю1ог!в); 31 —тройничный нерв (пегуив 1гщет1пив); 32 — вестибулярный нерв (пегуив уе511Ьи1ат). Стрелкой обозначен свод
Тема 3. Теорія функціональних систем П.К.Анохіна. Функціональна система. Стадії поведінкового акту. Поведінка у вирогідністному середовищі. Нейронні механізми поведінки. Функціональні стани та їх фізіологічні індикатори. Теория функциональных систем Что такое система? Термин «система» обычно применяется для того, чтобы указать на собранность, организованность группы элементов и ограниченность ее от других групп и элементов. Давалось множество определений системы, которые характеризовали ее, выделяли из «несистем». В общем, эти определения сводились к пониманию системы как комплекса взаимодействующих элементов, объединенных определенной структурой. При этом под структурой понимались законы связи и функционирования элементов. Однако понятие о «взаимодействии» элементов само по себе, с одной стороны, не дает исследователю в какой-либо конкретной области науки ничего нового, так как является даже для начинающего исследователя аксиомой. Кроме того, «взаимодействие» не объясняет, как ограничивается огромное число степеней свободы каждого из множества элементов живых систем. А без такого ограничения, упорядочивающего организацию элементов, живая система не возникает. Их взаимодействие часто создаст не упорядоченность, а хаос. Главным препятствием для использования в конкретном исследовании разнообразных вариантов системного подхода, базирующихся на указанном выше понимании системы, является отсутствие в их методологии понятия о системообразующем факторе, детерминирующем формирование и реализацию системы. До тех пор, пока исследовательно определит такой фактор, который: а) являясь неотъемлемым компонентом системы, ограничивал бы степени свободы ее элементов, создавая упорядоченную организацию, и б) был бы изоморфным для всех систем, позволяя использовать систему как инструмент анализа в самых разных ситуациях, этому исследователю будет сложно обнаружить преимущества системного подхода перед несистемным. В. Гейзенберг подчеркивает, что в соответствии с квантовой теорией нет никакого предшествующего события, из которого с необходимостью должно в определенное время последовать другое событие, например излучение альфа-частицы атомом радия. В то же время, если рассматривать живой организм не как целостного индивида, совершающего приспособительное поведение, а как физическое тело, то представление о детерминации внешней причиной — стимулом — может быть удобным приближением, справедливым в рамках этой ограниченной области описания. Разные типы детерминации связаны, и вся реальность несводима к единственному типу детерминации. Однако представление о телеологической, целевой детерминации может быть использовано лишь для описания живого. Именно с этой детерминацией связана специфика процессов, изучаемых психологией, психофизиологией, биологией. И именно по критерию «активность — целенаправленность — реактивность» могут быть наиболее четко дифференцированы представления системной и традиционной психофизиологии и нейронауки. Поэтому противопоставление телеологической и стимульной детерминаций представляется дидактически оправданным. Определение эмоции Что такое эмоция? Уильям Джемс — автор одной из первых физиологических теорий эмоций — дал своей статье, опубликованной более 100 лет тому назад, весьма выразительное название: «Что такое эмоция?». Однако и через 100 лет после того, как был сформулирован этот вопрос, мы можем прочитать в руководстве «Физиология человека» следующее признание: «Несмотря на то что каждый из нас знает, что такое эмоция, дать этому состоянию точное научное определение невозможно... В настоящее время не существует единой общепризнанной научной теории эмоций, а также точных данных о том, в каких центрах и каким образом эти эмоции возникают и каков их нервный субстрат». С этим положением руководства можно согласиться и сегодня. Большинство исследователей рассматривает эмоцию как сложный психофизиологический феномен, который можно связать с: 1) переживаемым и/или осознаваемым чувством (состоянием) — феноменология эмоций, 2) висцеральными процессами, сопровождающими эмоции; 3) выразительными характеристиками эмоций (экспрессия лица, интонация, жесты и позы). Понимание сущности всякого психического процесса невозможно без знания мозговых механизмов, поэтому в психофизиологии эмоций необходимо выделить еще один важный аспект — центральные нейронные механизмы эмоций. При рассмотрении нейронных механизмов исследователи обращают основное внимание на устройство нейронной сети, связанной с тем или иным психическим процессом, и на функционирование этой сети, которое выражается в электрической активности отдельных нервных клеток. Функционирование нервной сети невозможно без связей между нейронами, которые осуществляются через синапсы, где главную роль выполняют специальные химические соединения медиаторы. Медиаторы участвуют в работе всех нервных сетей мозга, включая нейронные сети, связанные с эмоциями. Поэтому в современной психофизиологии эмоций выделяют еще один аспект - нейрохимические механизмы эмоций. Единицы памяти 'Не только окружающий мир, по и внутренний мир человека образован комбинациями из небольшого числа элементов. Если единицами окружающего мира служат атомы, молекулы, а единицами живой материи — гены, клетки, то простейшей физической единицей субъективного мира человека служит пакет (иначе — совокупность, кластер или группа) взаимосвязанных волн нейронной активности, порождаемых нервными клетками. О волновой природе единиц памяти впервые заговорил знаменитый английский врач Дэвид Гартли, основоположник психофизиологии. Пакеты нейронных волн генерируются нервными клетками, хотя и расположенными в разных центральных структурах, но взаимосвязанными, объединенными в ансамбли. Часть нейронов каждого такого ансамбля размещена в филогенетически древних структурах, в ретикулярной формации ствола мозга, в промежуточном мозге, в старой коре. Другие нейроны ансамбля размещаются в более дифференцированных колонках новой коры, в первичных-проекционных полях и в ассоциативных зонах. Первоначально нейроны объединяются в какой-либо ансамбль под влиянием воспринимаемого стимула, сохраняя в паттернах своей активности информацию о породившем их стимуле. Согласованная, периодически повторяющаяся активность нейронов в составе ансамбля необходима для поддержания их собственной жизнедеятельности. Повторяющаяся активность нейронов приводит со временем к структурным изменениям межнейронных связей и поддерживает устойчивость структурных изменений в дальнейшем. Подчеркнем, что изменения проводимости синапсов, т. е. контактов между нейронами, образующих так называемые векторы проводимости, всегда вторичны. Единичная волна в составе пакета связана с отдельной группой последовательных нейронных импульсов. Обычное число импульсов в группе — не более десяти на отрезке времени не больше десятой доли секунды. В известных опытах Н. П. Бехтеревой обнаружена связь подобных импульсных последовательностей с физическими и смысловыми особенностями воспринимаемых стимулов. Несколько последовательных волн, порождаемых нейронным ансамблем за один период, в свою очередь образуют пакет, кластер. Волны в пакете различаются своими фазами. Вероятно, первые по порядку волны в пакете возникают снач ла в менее дифференцированных древних структурах мозга, а более поздние волны — в более дифференцированных новых структурах. Периоды колебаний согласованной активности нейронов, объединенных в ансамбли, имеют разную длительность. Образы памяти закодированы разночастотными колебаниями. О составе периодов можно судить по спектрам мощности ЭЭГ. У человека преобладают периоды длительностью около 0,1 с. известные как альфа-ритм, названные так первооткрывателем Гансом Бергером В спектре мощности заметен пик амплитуды на этой частоте - Р = 10 Гц. Это константа Бергера Рядом, через интервал около 1 Гц, можно заметить, как правило, еще один пик Видны также и другие пики в разных диапазонах частот, но обычно в состоянии покоя испытуемого, при закрытых глазах пик в диапазоне альфа-ритма отличается наибольшей высотой, т. е. мощностью. У человека альфа-ритм выражен в наибольшей степени. Устойчивость частоты этого ритма на протяжении многих лет жизни взрослого испытуемого - одна из давно известных нейрофизиологических закономерностей. Испытуемые различаются по частоте доминирующего альфа-ритма примерно так же, как они различаются по росту, в целом в пределах 8-13 Гц, чаще всего — в пределах 9-11 Гц. Волны с близкими по длительности периодами, образуя на спектре мощности частоты, разделенные провалом около 1 Гц в диапазоне альфа-ритма, создают феномен веретен альфа-ритма, длительностью около 1 с. Веретена представляют собой плавные подъемы и спады амплитуды регулярных колебаний ЭЭГ. В состоянии покоя человека типичная длительность одиночного альфа-веретена равна 1 с. Ступенчатость различий между соседними частотами на спектре мощности, объясняющая феномен альфа-веретен, является примечательной нейрофизиологической закономерностью, впервые установленной М. Н. Ливановым [1989]. Относительная величина ступеньки — R = 0',1 — равна отношению минимальной разности периодов к их средней величине. Она названа константой Ливанова.
Теории фильтра Толчком к построению первых моделей внимания были результаты экспериментов по дихотическому прослушиванию, в которых испытуемый должен избирательно слушать один из двух или нескольких одновременно читаемых текстов. Первые такие эксперименты были проведены в 1950-х гг. Е. Черри. Он приводит пример вечеринки с коктейлями, на которой человек может слушать собеседника и не слышать разговоры других участников. Это фокусирование внимания. Иногда этот человек слышит от других беседующих свое имя. Это разделение внимания. На основании этого примера Е. Черри ставит ряд вопросов. Как мы можем настроиться на определенный источник информации, оставаясь чувствительными к значимой информации от других источников? Как наше внимание может быть сфокусировано и разделено одновременно? С момента появления работ по дихотическому прослушиванию избирательные и произвольные аспекты внимания стали рассматривать с формальных позиций информационной теории. Согласно этим представлениям, нервная система представляет коммуникационный канал (т.е. канал связи!) с ограниченными перцептивными возможностями. Для выбора одного канала информации из множества параллельных каналов в нервной системе предполагалось наличие специальных фильтров. Первую теоретическую модель внимания, или модель фильтра, создал Д- Бродмент. Согласно его модели, информация от рецепторов поступает в центральную нервную систему через множество параллельных сенсорных каналов связи. Однако нервная система, несмотря на множество входов, может работать как одиночный коммуникационный канал с ограниченными возможностями. Поэтому на входах этого канала осуществляется операция селекции, т. е. выбирается определенная сенсорная информация. Селекция не совсем случайна. Селективный фильтр может быть настроен на принятие желаемой информации. Предполагается, что в мозге есть нейронная репрезентация информации, которая контролирует работу фильтра по физическим признакам, например по интенсивности, высоте и пространственной локализации звука. Для всей остальной информации фильтр блокирует входы, но эта невостребованная информация какое-то время хранится в блоке кратковременной памяти, который находится перед фильтром. Первоначально невостребованная информация из блока кратковременной памяти может пройти через коммуникационный канал, если произойдет сдвиг селективного процесса с одного класса сенсорных событий на другой. Схема Д. Бродмента не отвечает на вопрос, поставленный еще Е. Черри: почему мы переключаем внимание на какой-либо сенсорный вход, если не знаем, что на него поступило? Ответ на этот вопрос может быть получен в рамках модели А. Трейсман (Treisman). В ее модели постулируется двухстадийная фильтрация. Весь поток информации, как и в предшествующей модели, поступает в организм через множество параллельных каналов. На некотором уровне нервной системы находится 1-й фильтр, где происходит выделение по физическим свойствам (громкость, высота, позиция в пространстве, цвет, яркость и т. д.) одного из каналов, по которому сигналы проходят беспрепятственно, и одновременно происходит ослабление сигналов по другим каналам. Ослабленные и неослабленные сигналы проходят через 2-й фильтр — логический анализатор (словарь), который представлен специализированными нейронами. Активность каждого такого нейрона связана с определенным словом, составляющим словарь индивида, и приводит к осознанию субъектом слов. Эти нейроны активируются неослабленными сигналами, а некоторые из них с достаточно низким порогом чувствительности могут быть активированы и ослабленными сигналами. В модели Дж. Дойч и Д. Дойч фильтр в явной форме отсутствует. Согласно их модели, все сигналы доходят до логического анализатора, где каждый из них анализируется на предмет специфичности. Чем важнее сигнал для организма, тем более выражена активность нейронов логического анализатора, на которые он поступил, вне зависимости от его исходной силы. Важность сигнала оценивается на основе прошлого опыта. Работа логического анализатора не контролируется сознанием. Осознается только информация, выходящая из него. Теории вызванного внимания Общая идея теорий фильтра заключается в том, что на пути прохождения электрических импульсов (возбуждения) по нервным волокнам от рецепторов, подвергшихся воздействию внешних раздражителей, до коры имеет место механизм, отфильтровывающий эти импульсы. Нейро- и психофизиологи в своих исследованиях мозговых механизмов внимания взяли на вооружение эту идею. Многие экспериментальные исследования были направлены па изучение судьбы афферентных возбуждений, вызванных тем или иным сенсорным раздражением в ситуации привлечения внимания к этому раздражению или при отвлечении от него внимания. Исследователи рассуждали следующим образом. Существуют морфологически выделяемые афферентные пути - от рецепторов до коры головного мозга. Эти пути имеют синаптические переключения в ядрах ствола головного мозга и в коре. Существуют эфферентные пути от коры и ретикулярной формации ствола мозга к ядрам, где происходит переключение афферентных путей. Известно, что потенциалы действия или возбуждения, идущие по эфферентным путям, могут изменять синаптическую передачу афферентных возбуждений. Следовательно, процесс фильтрации в виде торможения возбуждений по некоторым афферентным волокнам происходит на этапах переключения этих путей в релейных ядрах. При этом допускалось, что показателем торможения является уменьшение импульсной активности нейронов этих ядер, что должно сопровождаться уменьшением их суммарной активности, регистрируемой в виде вызванных потенциалов (ВП). Несомненно, что из сказанного выше следует однозначный вывод: поскольку внимание проявляется через избирательное торможение в определенных сенсорных каналах, то, следовательно, животное заранее знает, что они не соответствуют ситуации. Результаты первых исследований подтвердили эти предположения. В известных экспериментах Р. Эрнандец-Пеона [1979] предъявления «незнакомого» звукового щелчка вызывали у кошки ориентировочное поведение в виде заинтересованности этими щелчками и большие по амплитуде вызванные потенциалы (ВП) в улиточном ядре. Дальнейшие однообразные предъявления этого щелчка приводили к угашению внимания к щелчкам (привыканию, или габитуации), что сопровождалось сначала исчезновением ориентировочного поведения (кошка засыпала) и лишь через длительный промежуток времени уменьшением амплитуды ВП вплоть до его исчезновения. Если звук щелчка, внимание к которому было угашено, внезапно изменяли (изменяли громкость, локализацию или тембр), то кошка пробуждалась (дисгабитуация), а в корковых и в подкорковых областях увеличивалась амплитуда ВП. В других опытах Р. Эрнандец-Пеоном и его последователями было показано, что ВП на сенсорные сигналы в коре и подкорковых образованиях уменьшаются и У кошек, и у людей по амплитуде при отвлечении внимания от этих сигналов. Отвлекающим фактором для кошек могли быть запах пищи, появление в поле зрения мыши или крысы, писк крысы, электрокожное раздраженние. У человека также уменьшалась амплитуда ВП в слуховой коре, если внимание от раздражителей отвлекали тем, что он решал какие-либо задачи, читал или вспоминал события своей жизни. Казалось, что проблема фильтров близка к разрешению - повышение внимания к сигналу сопровождается усилением связанного с ним афферентного потока, и, наоборот, угашение внимания к сигналу ослабляет этот поток. I выдвинуто предположение, что угашение и отвлечение внимания связаны с активностью ретикулярной формации, участвующей в блокаде афферентных возбуждений. Однако в других лабораториях, где проводились аналогичные исследования не было обнаружено каких-либо закономерных изменении амплитуды ВП в слуховых путях на щелчки, к которым предварительно было угашено внимание. Так у кошек в правом и левом слуховых ядрах и даже в разных точках одного ядра исследователи наблюдали разнонаправленные изменения амплитуды ВП, неоднозначные изменения наблюдались также в амплитуде корковых ВП. Более того, как показали исследования Л. Г. Воронина и Е. Н. Соколова если интенсивность сигнала, к которому было выработано привыкание (т. е. фильтры не должны пропускать от него афферентное возбуждение), внезапно снизить до порогового уровня то наблюдается дисгабитуация со всеми ее проявлениями. На основании этих данных уже невозможно утверждать, что невнимание к сигналу осуществляется через блокаду афферентных возбуждений где-то на этапах их синаптических переключений. Вместо него было выдвинуто предположение о том, что все афферентные возбуждения поступают в центр. На последнем предположении построена теория нервной модели стимула Е. Н. Соколова. Согласно этой теории, в процессе привыкания к внешним раздражителям в коре формируется нервная модель стимула, в которой фиксированы все параметры знакомого комплекса раздражителей. Такая модель, по мнению Е. Н. Соколова, обеспечивает высокий уровень внимания, поскольку позволяет воспринимать информацию от рецепторов, возбужденных знакомыми раздражителями, за более короткое время и более надежно, чем возбуждения от малознакомых раздражителей. Возбуждения, поступившие в центр от незнакомого раздражителя, оказываются несогласованными с существующей нервной моделью, в результате чего формируется ориентировочное поведение, которое можно рассматривать как внимание, направленное на этот раздражитель. Развивая теорию о нервной модели стимула, Р. Наатанен предположил возможность формирования трех различных типов нервных моделей стимула. Одна из них — «пассивная», или непроизвольная, модель стимула — формируется после многократного предъявления стимула («стандартный» стимул) даже при отвлечении от него внимания и сохраняется в течение примерно 5 с. Если в пределах этого интервала предъявляется «отклоняющийся» стимул, который отличается по физическим характеристикам от часто предъявляемого «стандартного» стимула, то происходит рассогласование приходящих от этого «отклоняющегося» стимула афферентных возбуждений с нервной моделью «стандартного» стимула. В электрической активности мозга, а именно в связанных с событием потенциалах ( ССП), это рассогласование проявляется в виде негативной волны, получившей название негативность рассогласования (НР). НР развивается через 100 мс после предъявления стимула и длится около 250 мс. Чем больше разница между «стандартным» и «отклоняющимся» стимулами, тем больше амплитуда НР. Процесс рассогласования не осознается, но он может привлечь внимание субъекта к изменениям в последовательности предъявляемых стимулов. Предполагается, что НР является необходимым, но не достаточным условием для осознанного восприятия изменении в стимуле. В случаях привлечения внимания к стимулу, например, когда испытуемый должен считать какой-то редкий стимул на фоне более частых, у него формируется «активная», или произвольная, нервная модель часто предъявляемого стимула. Время существования этой модели определяется тем, насколько долго испытуемый сохраняет внимание к выполняемой задаче. Данная модель является своего рода стандартом, с которым сравниваются поступающие от стимулов афферентные возбуждения. Афферентные возбуждения от редкого сигнала вызывают процесс рассогласования, который и обеспечивает узнавание редкого сигнала. В ССП процесс рассогласования с «активной» моделью сопровождался негативным колебанием Н-2, которое состояло из НР и дополнительного негативного колебания Н-26. Чем больше выражены рассогласования афферентных возбуждений от стимула с пассивной или активной моделями, том больше амплитуда соответствующих волн. На основании этих данных авторы считают, что пассивная и активная модели существуют одновременно и обеспечиваются активностью нейронов разных областей мозга. Когда вероятность появления стимула, который нужно обнаружить (его еще называют целевым стимулом), возрастает до определенного уровня, то уже формируется нервная модель именно этого стимула. Авторы называют эту модель «следом внимания», поскольку она формируется и поддерживается стимулом, к которому привлечено внимание. «След внимания» существует только тогда, когда у субъекта актуализирован ясный образ целевого стимула. Распознавание целевого стимула в этом случае происходит за счет процесса согласования поступающих от него афферентных возбуждений с его же нервной моделью. Развивающаяся во время этого процесса негативная волна, или негативность, по-видимому, связана с обработкой информации. Эта волна начинается на нисходящем фронте Н-1 и продолжается в течение нескольких сотен миллисекунд. Чем больше афферентные возбуждения от целевого стимула соответствуют «следу внимания», тем большую амплитуду и длительность имеет эта волна. Итак, в основе модели внимания Р. Наатанена лежит нервная модель стимула, которая представляет собой репрезентированный в нейронных системах образ определенного стимула. Сопоставление афферентных возбуждений от стимула с моделью осознанно или неосознанно ожидаемого стимула характеризует внимание и проявляется в негативном отклонении компонентов ССП. В настоящее время НР как показатель уровня внимания внедряется в клиническую практику. В других теориях внимания, разрабатываемых в рамках информационной парадигмы, внимание определяется как процесс, лежащий в основе селекции и организации доступной информации для соответствующего ответа. Авторы связывают последовательность этих процессов с последовательными компонентами ССП: ранние позитивные компоненты ССП отражают регистрацию и анализ приходящей информации; внимание, осуществляющее селекцию этой информации, отражается в развивающейся вслед за позитивным компонентом негативной волне; поздняя позитивная волна отражает процесс выбора ответа. Недостатки теорий вызванного внимания. В описанных выше информационных моделях и концепциях внимания в основном рассматривается зависимость афферентных возбуждений от внешних источников, внимание к которым, по условиям эксперимента, привлекается или, наоборот, отвлекается. При этом авторы учитывают в своих теориях экспериментально доказанные факты влияния на нейроны релейных ядер, где эти афферентные возбуждения переключаются, и на рецепторные образования эфферентных возбуждений от корковых, подкорковых и стволовых структур мозга. Именно эфферентные возбуждения из Центральных структур, отражая собой индивидуальный опыт, и обеспечивают работу фильтров. Эти теории рассматривают внимание как механизм, вызванный внешними источниками, которые, по сути, являются строго определенными звуками или зрительными изображениями (иногда иллюзиями), навязанными экспериментальной ситуацией, применительно к которой только и имели смысл получаемые результаты. Но совершенно очевидно, что объектом внимания могут быть не только эти источники. Уже У. Джемс в конце XIX в. выделял как внешние (объекты и события в окружающей среде), так и внутренние (память и знания) источники, к которым привлекается внимание. Однако внимание к внутренним источникам вряд ли можно объяснить, используя гипотетический механизм последовательной обработки информации. С помощью только этого механизма невозможно объяснить и другой важный факт, который долго игнорировался в когнитивной психологии, а именно: эффекты внимания определяются обучением и тренировкой. Неэффективное выполнение какой-либо задачи на внимание свидетельствует об отсутствии навыка в выполнении этой задачи. Так, известный психолог рей, исследовавший внимание и многократно участвовавший как испытуемый экспериментах по дихотическому слушанию, приобрел навык легко обращать две деятельности одновременно: это записывание слов под диктовку и чтение про себя.
Основные концепции сознания Говоря о мозговых основах психики и сознания, необходимо отметить, что эти наиболее сложные проявления работы мозга не могут быть объяснены работой какой-то отдельной, изолированной группы нервных клеток, представляющих собой «центр сознания». Как общетеоретические соображения, так и данные экспериментов свидетельствуют о том, что психические функции возникают на основе объединения в единую систему разных, функционально неоднородных нервных структур и специальной организации процессов мозга, обеспечивающей высокую степень мозговой интеграции. Изучение этой организации и лежит в основе поиска мозговых основ сознания. В настоящей главе не ставится задача изложить многие и весьма интересные данные о роли мозговых структур в осуществлении психических функций. Количество таких данных в последнее время стремительно увеличивается, особенно после появления методов «изображения живого мозга», таких как позитронно-эмиссионная томография, функциональный магнитный резонанс, компьютерная электроэнцефалография и др. «Светлое пятно» Гипотеза о связи сознания с определенным участком коры была впервые высказана И. П. Павловым в 1913 г. Хотя, как известно, психологические термины по принципиальным соображениям не использовались в лабораториях И. П. Павлова, он считал, что понимание мозговых основ сознания — одна из главных задач науки о мозге. В поисках ответа на вопрос, «какие нервные процессы происходят в больших полушариях тогда, когда мы говорим, что мы себя сознаем», он предположил, что сознание представлено деятельностью находящегося в состоянии оптимальной возбудимости «творческого» участка коры больших полушарий, где легко образуются условные рефлексы и дифференцировки. Другие участки, где происходит преимущественно поддержание уже образованных рефлексов, связаны с тем, что называется бессознательной деятельностью. Свои мысли И. П. Павлов выразил в образной форме: «Если бы можно было видеть сквозь черепную коробку и если бы место с оптимальной возбудимостью светилось, то мы увидели бы на думающем сознательном человеке, как по его большим полушариям передвигается постоянно изменяющееся в форме и величине причудливо меняющихся очертаний светлое пятно». И. П. Павлов, правда, оговаривался, что он не касается при этом «философской точки зрения», каким образом материя мозга производит субъективное явление. Фактически это то немногое, что сказал создатель учения о высшей нервной деятельности по проблеме сознания. Тем более удивительно, как это высказывание И. П. Павлова пророчески подтвердилось. Мы можем теперь действительно видеть «сквозь черепную коробку» активные зоны с помощью методов «изображения живого мозга», о которых говорилось ранее, и видим нечто похожее на то, о чем говорил И. П. Павлов. Главное же заключается в том, что концепция «светлого пятна» получила свое развитие в новейших гипотезах в виде «теории прожектора». Данная концепция была сформулирована Ф. Криком - соавтором теории двойной спирали и нобелевским лауреатом. Здесь хотелось бы сделать небольшое отступление: можно сказать, что проблемы сознания как бы притягивают к себе внимание нобелевских лауреатов, хотя никто еще Нобелевскую премию за исследования сознания пока не получил. И. П. Павлов был только первым, но не последним в этом ряду, ведь проблема сознания — одна из важнейших и интереснейших в науке, и лучшие умы, воодушевленные высоким признанием, тянутся к ее разрешению. Основные предпосылки и положения «теории прожектора» сводятся к следующему. Вся информация поступает в кору по сенсорным путям через переключательные ядра в дорзальном таламусе (включая и примыкающие к нему коленчатые тела). Возбудимость этих переключательных ядер может быть избирательно изменена за счет коллатералей от нейронов ретикулярного комплекса таламуса, входящего в его вентральный отдел. Взаимоотношения между этими частями таламуса построены таким образом, что в каждый данный момент одна из нейронных групп дорзального таламуса оказывается в состоянии высокой возбудимости, что значительно усиливает импульсный поток к коре, в то время как другие группы оказываются, наоборот, заторможенными. Период такой высокой возбудимости длится около 100 мс, а затем усиленный приток поступает к другому отделу коры. Ф. Крик предполагает на этом основании, что область наиболее высокой импульсации представляет в данный момент как бы центр внимания, а благодаря перемещению «прожектора» в другие участки становится возможным их объединение в единую систему. В этом процессе большую роль играет синхронизация активности нейронных групп на одной частоте. В последние годы особое значение в процессах интеграции придается высокочастотному ритму — частотой около 40 Гц. Важно, что в число вовлеченных в совместную деятельность нейронных групп входят нейронные ансамбли в различных областях коры. Подобная интеграция, по мнению автора, и обеспечивает осуществление высших психических функций. Предполагается также, что одновременно могут действовать несколько «прожекторов». Данная концепция была разработана, в первую очередь, для обработки зрительных сигналов, но автор предполагает, что описанная схема является общей для всех анализаторов. Сознание, общение и речь. Концепция коммуникативной природы сознания была глубоко разработана П. В. Симоновым. По его определению, сознание представляет собой знание, которое в абстрактной форме может быть передано другим людям, что находит отражение в этимологии слова «сознание» — «совместное знание». Сознание возникло в процессе эволюции на базе потребности к общению, передачи знаний и объединения усилий высокоорганизованных членов сообщества, какими являлись наши предки. Однако поскольку внутренний мир человека скрыт от внешнего наблюдателя, передача сведений от одного человека к другому может происходить лишь путем абстракции, т. е. в виде знаков. Такой знаковой формой общения является речь, формирующаяся в процессе общения. На основе общения возникает и сознание как высшая форма Сознание как высшая форма психического возникает на основе общения между людьми. психического, свойственная только людям. Непосредственная связь сознания с речью показана в исследованиях людей, выходящих из состояния комы. В этом случае речевой контакт с больным, что, как известно клиницистам, является важным признаком возвращения сознания, совпадает с образованием когерентных связей между электрической активностью гностических (теменно-височных) и моторно-речевых (нижнелобных) отделов левого полушария. Э. А. Костандов считает передачу сигналов на моторные речевые центры решающим условием для перехода от бессознательных к осознаваемым формам восприятия внешних сигналов Особая роль левого, речевого полушария в механизмах сознания была показана и в исследованиях больных с перерезкой мозолистого тела (нервный пучок, соединяющий полушария). Было установлено, что в первые недели после операции у подо   Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ТАКОЕ УВЕРЕННОЕ ПОВЕДЕНИЕ В МЕЖЛИЧНОСТНЫХ ОТНОШЕНИЯХ? Исторически существует три основных модели различий, существующих между...  ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|