
|
|
Распределение заряда в молекуле ⇐ ПредыдущаяСтр 6 из 6 Все белки построены из 20 типов аминокислотных остатков; в зависимости от того, может ли боковая цепь данного остатка ионизовываться, его относят к одному из трех типов: кислотному (может иметь отрицательный заряд), основному (может иметь положительный заряд) и нейтральному [ВОЛЬКЕНШТЕЙН, 1988]. Есть три типа кислотных остатков (Asp, Glu, Tyr) и три типа основных остатков (His, Lys, Arg). Заряд аминокислотного остатка зависит от pH растворителя, в который он погружен. При pH=7.0, соответствующем естественному окружению фермента, ионизованы только четыре остатка: Asp, Glu, Lys и Arg. Все дальнейшие вычисления производятся для pH=7.0. Таким образом, мы знаем заряды и координаты всех ионизированных атомов белка и можем использовать их для вычисления электростатических характеристик фермента. Прежде чем перейти к полученным результатам, необходимо сказать о том, каким образом используемая система координат ‘привязана’ к молекуле фермента. За начало отсчета принят атом CD1 остатка На рис. 6.5 представлена интегральная плотность заряда вдоль оси Oz, т. е. вдоль оси ущелья. Кривая показывает суммарный заряд между плоскостями, перпендикулярными оси ущелья; одна из плоскостей фиксирована и пересекает Oz в нуле.
Рис. 6.5. Интегральная плотность распределения заряда фермента вдоль оси Oz в единицах заряда электрона.
Заряд «верхней» половины фермента отрицателен и составляет -14e, заряд «нижней» половины равен +2e. Таким образом, АХЭ имеет отрицательный заряд -12e. Число ионизованных атомов равно 110. Ассиметрия распределения заряда показывает, что молекула должна обладать дипольным моментом. В дальнейшем мы определим дипольный момент при различных ионных силах растворителя для сравнения с экспериментальными данными. Совпадение результатов косвенно указывало бы на правильность рассчитанного потенциала, к вычислению которого мы сейчас переходим.
6.3.2 Расчет потенциала В естественных условиях молекулу АХЭ окружают различные ионы, содержащиеся в интерстициальной жидкости. Поэтому при расчете потенциала в растворителе необходимо учитывать влияние ионной атмосферы, возникающей вокруг каждого заряда и изменяющей его поле. Действительно, пусть имеется растворитель, содержащий различные сорта ионов. Обозначим их валентности через
где
Плотность заряда в данной точке растворителя получается суммированием плотностей заряда, даваемых каждым сортом ионов:
Поскольку исходный растворитель электрически нейтрален, первая сумма обращается в нуль. Тогда
где I ¾ ионная сила раствора,
В дальнейшем будет видно, что соотношение
Табл. 6.1. Ионный состав интерстициальной жидкости.
Используемая здесь единица концентрации M соответствует одному молю растворенного вещества в одном литре раствора: 1 M = 1 моль/литр. Итак, считая наведенную ионами плотность заряда равной
где
Уравнение Пуассона для потенциала примет вид
где Данное уравнение числено решалось на сетке 100´100´100 точек, шаг сетки соответствовал 1 На рис 6.6 изображены эквипотенциали электростатического поля при нулевой ионной силе. Указанные энергии соответствуют заряду +1e. На рис. 6.7 представлен потенциал фермента на оси ущелья при ионных силах 0, 10 и 20 мМ.
а) б) Рис. 6.6. Эквипотенциали электростатического поля молекулы АХЭ при нулевой ионной силе. Указанные энергии соответствуют заряду +1e. а) «Черный ход» закрыт; б) «черный ход» открыт.
Рис. 6.7. Потенциал АХЭ на оси ущелья активного центра
при различных ионных силах. Возвращаясь к вопросу о применимости теории Дебая-Хюккеля видим, что в точке с наименьшим потенциалом
6.3.3 Определение дипольного момента Зная теперь не только заряды атомов белка, но и плотность наведенного заряда, можем вычислить дипольный момент системы фермент плюс ионная атмосфера:
где
где
Рис. 6.8. Зависимость дипольного момента d от ионной силы I. Квадратиками показаны рассчитанные значения дипольного момента; кружками - экспериментальные.
Математическая модель. Используя результаты рентгеноструктурного анализа, представим фермент набором жестко связанных неподвижных точек, относительное расположение которых соответствует расположению атомов в молекуле AChE. Для сравнения скорости выхода “осколков” из ущелья в случае закрытого и открытого “back door” используем две модели фермента. Первая модель соответствует молекуле AChE с закрытой “back door” и содержит все атомы, определенные рентгеноструктурным анализом. Вторая модель, соответствующая ферменту с открытой “back door”, должна иметь канал, через который продукты реакции могли бы покинуть ущелье активного центра. Поэтому из второй модели необходимо исключить некоторое количество атомов, определенных рентгеноструктурным анализом. Сделаем это следующим образом. Будем считать, что канал имеет цилиндрическую форму, радиус канала примем равным 5 Субстрат и его “осколки” также представим набором жестко связанных точек. Однако, в данном случае для уменьшения времени счета не будем считать подвижные объекты состоящими из большого числа атомов, а отобразим молекулы ацетилхолина и холина в виде гантелей и молекулу ацетата в виде шара. Размеры гантелей и шара определим из результатов рентгеноструктурного анализа для соответствующих молекул. Наделим каждую точку соответствующими массой и зарядом. Ван-дер-ваальсово взаимодействие двух точек, принадлежащих различным объектам (например, ферменту и продукту реакции), представим потенциалом Леннарда-Джонса:
где r - расстояние между точками; константы A и B для различных пар атомов приведены в [1]. Учет ударов молекул растворителя произведем по методу Ланжевена, т. е. введем в уравнения движения случайную силу [17]. Полагая фермент жестко закрепленным, будем описывать трансляционное движение подвижных объектов координатами и скоростями их центров масс. Для описания вращательного движения с центром масс каждого объекта свяжем подвижную систему координат. Ориентируем ее оси таким образом, чтобы тензор инерции принял диагональный вид. Это можно сделать следующим способом. В системе координат, связанной с центром масс, рассчитывается тензор инерции, определяются его собственные вектора и оси новой системы направляются вдоль собственных векторов. Тогда уравнения движения l-го тела запишутся наиболее просто [18]:
Здесь ml - масса тела; xl, yl, zl - координаты центра масс в декартовой системе; Jl1, Jl2, Jl3 - компоненты тензора инерции; Wl1, Wl2, Wl3 - частоты вращений вокруг осей собственной системы координат;
xlx, xly, xlz - проекции случайной силы на оси лабораторной системы координат; zl1, zl2, zl3 - проекции случайного момента сил на оси подвижной системы координат; gl - коэффициент, учитывающий силу вязкого трения; gl1, gl2, gl3 - коэффициенты, учитывающие силу вязкого трения при вращательном движении;
Сила
Здесь В процессе численного счета трансляционное брауново движение моделировалось следующим образом. Через равные интервалы времени Аналогичным образом моделировалось вращательное брауново движение. Через равные интервалы времени В качестве примера рассмотрим определение скорости связывания субстрата с ферментом или константы k1.
§3.4. Определение константы скорости k1.
Константа k1 определяет скорость образования комплекса E^S из независимых молекул фермента и субстрата: w1=k1[E][S]. Воспользуемся следующей схемой определения этой константы. Ограничим область вблизи основного входа в ущелье активного центра непроницаемыми для молекулы субстрата стенками в форме цилиндра (рис. 3.5). Поместим случайным образом в эту область молекулу ACh и промоделируем ее диффузию в поле, порождаемом ферментом. Если z-компонента координат субстрата становится меньше z1, то считаем комплекс E^S образовавшимся. Многократно повторяя данный этап, определим частоту n образования комплекса E^S при условии, что молекула субстрата все время находится в данной области.
Рис. 3.5. Объем, ограничивающий область около входа в активный центр. Субстрат расположен на оси Oz в точке z1.
Пусть средняя по объему концентрация субстрата в выделенной области равняется [S]1. Тогда среднее отношения времени t, когда субстрат находится в выделенной области, ко времени наблюдения T равняется
где V1 - объем выделенной области. Записанное соотношение справедливо при условии [S]1V1<<1; иначе необходимо принимать во внимание те моменты времени, когда в объеме V1 находятся две и более молекул субстрата. Очевидно, при наличии только одной свободной молекулы фермента частота образования комплекса E^S равняется k1[S]; следовательно,
где q - заряд субстрата (+1e). Введем обозначение
тогда
В процессе численного счета объем V1 ограничивался плоскостью z2=35
Рис. 3.6. Зависимость количества образовавшихся E^S-комплексов N от времени моделирования при нулевой ионной силе.
Рис. 3.7. Зависимость константы скорости k1 от ионной силы I. Указанные погрешности соответствуют 66%-му доверительному интервалу.
РЕЗУЛЬТАТ МОДЕЛИРОВАНИЯ С «BACK DOOR»
Рассматривалась возможность объяснения эффекта субстратного торможения увеличением времени выхода продуктов реакции из ущелья. Это рассмотрение могло бы дать ответ на вопрос о том, имеется ли “back door” в молекуле AChE. Действительно, если бы рассчитанное значение [S]опт для молекулы фермента без “back door” оказалось значительно меньше экспериментального, мы имели бы аргумент в пользу наличия дополнительного канала в ущелье активного центра. Однако даже при концентрации субстрата, на порядок превосходящей экспериментальное значение [S]опт, среднее время выхода оставалось на четыре порядка меньше времени, определяющего химические стадии реакции. Таким образом, на основании имеющихся экспериментальных данных мы не можем утверждать, что в молекуле AChE происходит открытие дополнительного канала в ущелье активного центра.
ЗАКЛЮЧЕНИЕ
Таким образом мы рассмотрели математическую модель связывания субстрата. С моделями других этапов работы молекулярных ножниц можно ознакомиться по монографиям [1,2].
  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право...  ЧТО ПРОИСХОДИТ ВО ВЗРОСЛОЙ ЖИЗНИ? Если вы все еще «неправильно» связаны с матерью, вы избегаете отделения и независимого взрослого существования...  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
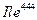 (Ile444.CD1), лежащий в основании ущелья активного центра. Ось Oz проходит через центр входа в активный участок и является, таким образом, осью ущелья. Центр входа в активный участок определен как среднее координат Glu73.CA, Asn280.CB, Asp285.CG и Leu333.O. Оси Ox и Oy ориентированы так, что атом Glu73.CA лежит в плоскости Oyz. Z -компонента координат центра входа и y-компонента координат атома Glu73.CA положительны, система координат является правой.
(Ile444.CD1), лежащий в основании ущелья активного центра. Ось Oz проходит через центр входа в активный участок и является, таким образом, осью ущелья. Центр входа в активный участок определен как среднее координат Glu73.CA, Asn280.CB, Asp285.CG и Leu333.O. Оси Ox и Oy ориентированы так, что атом Glu73.CA лежит в плоскости Oyz. Z -компонента координат центра входа и y-компонента координат атома Glu73.CA положительны, система координат является правой.

 , концентрации ¾ через
, концентрации ¾ через  . При помещении в растворитель какой-либо системы зарядов концентрации ионов
. При помещении в растворитель какой-либо системы зарядов концентрации ионов  перестанут быть однородными в пространстве. Согласно распределению Больцмана
перестанут быть однородными в пространстве. Согласно распределению Больцмана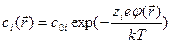 ,
, ¾ электростатический потенциал. В широко распространенной теории Дебая-Хюккеля предполагается, что справедливо соотношение
¾ электростатический потенциал. В широко распространенной теории Дебая-Хюккеля предполагается, что справедливо соотношение  . Тогда, разлагая экспоненту в ряд и пренебрегая членами второго порядка малости, запишем:
. Тогда, разлагая экспоненту в ряд и пренебрегая членами второго порядка малости, запишем: .
. .
. ,
, .
. ,
,  ,
,  и
и  (табл. 6.1).
(табл. 6.1).


 ,
, ¾ концентрация положительных одновалентных ионов,
¾ концентрация положительных одновалентных ионов,  ¾ концентрация отрицательных одновалентных ионов,
¾ концентрация отрицательных одновалентных ионов,  из-за нейтральности раствора, получаем:
из-за нейтральности раствора, получаем: .
. ,
,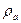 - плотность распределения зарядов атомов фермента.
- плотность распределения зарядов атомов фермента. , оператор Лапласа аппроксимировался разностным оператором, полученная система решалась методом простых итераций. Значение диэлектрической проницаемости принято равным 80.
, оператор Лапласа аппроксимировался разностным оператором, полученная система решалась методом простых итераций. Значение диэлектрической проницаемости принято равным 80.

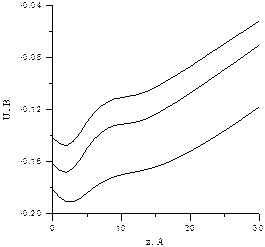
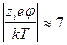 и ее использование в данном случае недопустимо.
и ее использование в данном случае недопустимо. ,
, в случае растворителя означает произведение плотности заряда в данной точке на объем ячейки дискретизации. Дипольный момент системы зарядов, сумма которых отлична от нуля, зависит от выбора системы координат, поэтому расчет производился относительно центра масс по формуле
в случае растворителя означает произведение плотности заряда в данной точке на объем ячейки дискретизации. Дипольный момент системы зарядов, сумма которых отлична от нуля, зависит от выбора системы координат, поэтому расчет производился относительно центра масс по формуле ,
,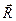 - радиус-вектор центра масс. Результаты вычислений для некоторых значений ионной силы вместе с экспериментальными данными, основанными на измерении постоянного и переменного электрического дихроизма в полях различной силы, представлены на рис. 6.8. (Подробности см. в [1,2].
- радиус-вектор центра масс. Результаты вычислений для некоторых значений ионной силы вместе с экспериментальными данными, основанными на измерении постоянного и переменного электрического дихроизма в полях различной силы, представлены на рис. 6.8. (Подробности см. в [1,2].
 . Путем варьирования направляющего вектора оси цилиндра и координат некоторой точки, лежащей на оси “back door”, определим такое положение канала, при котором в него попадает наименьшее число атомов фермента. Удалив оказавшиеся внутри канала атомы (принадлежащие остаткам Trp84 и Met83, что согласуется с приведенными в [9] данными), получим вторую модель фермента.
. Путем варьирования направляющего вектора оси цилиндра и координат некоторой точки, лежащей на оси “back door”, определим такое положение канала, при котором в него попадает наименьшее число атомов фермента. Удалив оказавшиеся внутри канала атомы (принадлежащие остаткам Trp84 и Met83, что согласуется с приведенными в [9] данными), получим вторую модель фермента.
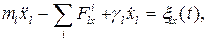
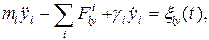




 ,
,  ,
,  - координаты i-ой точки тела в собственной системе координат;
- координаты i-ой точки тела в собственной системе координат; ,
,  ,
,  - проекции силы
- проекции силы  , приложенной к i-ой точке тела, на оси лабораторной системы;
, приложенной к i-ой точке тела, на оси лабораторной системы; ,
,  ,
,  - то же на оси подвижной системы координат.
- то же на оси подвижной системы координат.
 - ван-дер-ваальсова сила, действующая на i-ю точку l-го тела со стороны j-ой точки другого тела (индекс j пробегает по всем точкам всех тел, за исключением l-го тела);
- ван-дер-ваальсова сила, действующая на i-ю точку l-го тела со стороны j-ой точки другого тела (индекс j пробегает по всем точкам всех тел, за исключением l-го тела); 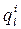 - заряд i-ой точки l-го тела;
- заряд i-ой точки l-го тела;  - напряженность поля.
- напряженность поля. скорость центра масс l-го тела изменялась на величину
скорость центра масс l-го тела изменялась на величину 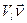 , где
, где  - амплитуда добавочной скорости,
- амплитуда добавочной скорости,  - равномерно распределенная в кубе [-1,1;-1,1;-1,1] случайная векторная величина. В остальное время тело двигалось согласно уравнениям (1) с нулевой случайной силой. Используя теорему о равнораспределении энергии по степеням свободы при диффузии в свободном пространстве в отсутствие электростатических полей
- равномерно распределенная в кубе [-1,1;-1,1;-1,1] случайная векторная величина. В остальное время тело двигалось согласно уравнениям (1) с нулевой случайной силой. Используя теорему о равнораспределении энергии по степеням свободы при диффузии в свободном пространстве в отсутствие электростатических полей 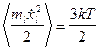 , получим следующее выражение для амплитуды шума:
, получим следующее выражение для амплитуды шума: 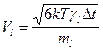 .
. , где
, где  - амплитуда добавочной угловой скорости, hi - равномерно распределенная на отрезке [-1,1] случайная величина. Амплитуды добавочных угловых скоростей задаются следующей формулой:
- амплитуда добавочной угловой скорости, hi - равномерно распределенная на отрезке [-1,1] случайная величина. Амплитуды добавочных угловых скоростей задаются следующей формулой:  .
.
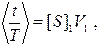

 Средняя концентрация [S]1 субстрата в области V1 выражается из распределения Больцмана следующим образом:
Средняя концентрация [S]1 субстрата в области V1 выражается из распределения Больцмана следующим образом: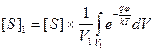 , (3.9)
, (3.9) ;
; . (3.10)
. (3.10)
