|
ГЛАВА 3. НАПРАВЛЕНИЯ МАКРОЭВОЛЮЦИИ И ПРОБЛЕМА ЕЕ НАПРАВЛЕННОСТИ
Проблема направленности эволюционного процесса остается в современной науке предметом оживленных дискуссий, крайние позиции в которых занимают, с одной стороны, сторонники жестко запрограммированной ("номогенетической") направленности эволюции (Л.С.Берг, А.А.Любищев, Л.И.Корочкин и др.), с другой же - ученые, разделяющие редукционистские взгляды об "оппортунистической эволюции", вообще лишенной какой-либо направленности (Д.Симпсон, Б.Ренш, Г.Стеббинс и др.). Чтобы в дальнейшем избежать путаницы, сразу же подчеркнем различие понятий направленности эволюции как закономерного проявления в макрофилогенезе некоторых общих тенденций, определенным образом организующих эволюционный процесс, и ее конкретных направлений в разных филетических линиях.
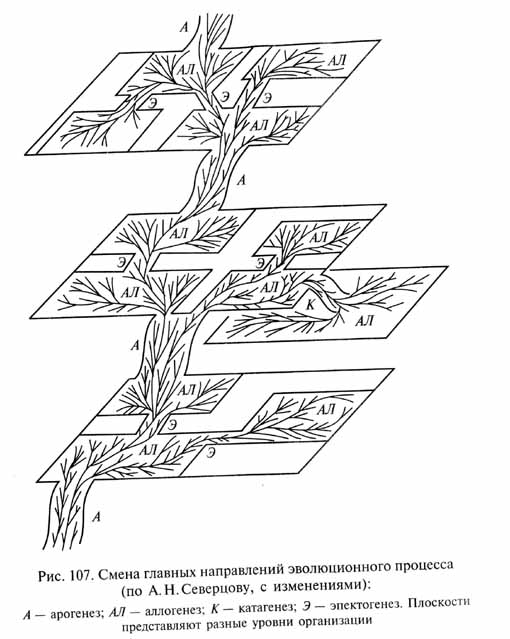
Главные направления эволюционного процесса А.Н.Северцов выделил три главных направления эволюционных преобразований: 1) морфофизиологический прогресс - повышение общего уровня организации, ее усложнение; 2) морфо-физиологический регресс (противоположное направление) - понижение и упрощение общего уровня организации; 3) идиоадаптация - развитие частных приспособлений, не изменяющих общий уровень организации. Позднее для обозначения этих направлений эволюции разными авторами были предложены другие наименования. Некоторые из них укоренились благодаря большей краткости и унификации звучания. Эволюционные преобразования организмов, ведущие к морфофизиологическому прогрессу и имеющие важное общее значение для организма в целом, А.Н.Северцов назвал ароморфозами. По аналогии с этим И.И.Шмальгаузен предложил называть регрессивные изменения катаморфозами, а термин "идиоадаптации" заменить на алломорфозы. В соответствии с этими терминами были предложены (А.Л.Тахтаджяном, А.А.Парамоновым и Б. Реншем) новые названия трех главных направлений эволюционного процесса: морфофизиологический прогресс стали именовать орогенезом (или анагенезом), морфофизиологический регресс - катагенезом, развитие частных приспособлений - аллогенезом (или кладогенезом).
(Еще одним особым направлением эволюции А.Н.Северцов первоначально считал ценогенезы (зародышевые приспособления), но позднее он сам признал это неоправданным, так как различные ценогенезы соответствуют тому или другому из других главных направлений эволюции).
Примеры указанных главных направлений эволюции можно найти в филогенезе основных групп хордовых животных. В целом по пути морфофизиологического прогресса шла эволюция подтипа позвоночных. Оболочники - группа, несомненно процветающая и в наше время, претерпели морфофизиологический регресс в связи с переходом их предков к прикрепленному образу жизни. Развитие идиоадаптации - всевозможных частных приспособлений - характеризует эволюцию любого крупного таксона (например, разнообразнейшие формы клюва у птиц, разные варианты строения конечностей у млекопитающих и т.п.). Многие группы длительное время эволюционировали именно путем идиоадаптаций, не изменяя общего уровня организации (например, любой из отрядов современных земноводных или пресмыкающихся, такие отряды млекопитающих, как грызуны, насекомоядные и т.п.). Понятие идиоадаптации (или алломорфозов) в концепции А.Н.Северцова объединяет очень широкий круг эволюционных изменений организмов - от самых незначительных частных приспособлений к специфическим условиям существования и образу жизни отдельных видов до адаптации общего значения, создающих предпосылки для значительного расширения среды обитания или освоения качественно новой адаптивной зоны (обширного круга местообитаний, нового способа использования ресурсов среды). Примерами таких "частных адаптации общего значения" являются кожное дыхание у земноводных, раковина моллюсков, особенности осевого скелета и мускулатуры тела змей, связанные с особым способом передвижения этих пресмыкающихся. Удачным примером подобного приспособления является усиленное развитие кутикулы в покровах нематод. Гибкая, прочная и химически устойчивая кутикула позволяет этим червям освоить широкий круг разнообразных местообитаний, включающих толщу органических осадков на дне морей и пресных водоемов, почву, эндопаразитизм в растениях и животных. Сильное развитие кутикулы стало отличительной чертой класса нематод и оказало существенное влияние на самые различные особенности их организации: оно обусловило редукцию мерцательного эпителия, утрату кольцевых мышц в кожно-мышечном мешке (поскольку прочная кутикула затрудняет перистальтические червеобразные движения); соответственно изменился характер передвижения нематод, вместо перистальтики возникли боковые изгибания тела; редукция же мерцательного эпителия сделала необходимым развитие нового типа органов выделения - нефридии заменились на шейные железы'.
Очевидно, эволюционное приобретение таких адаптации общего значения не соответствует ни собственно идиоадаптациям - частным приспособлениям, не оказывающим широкого влияния на организм в целом, ни ароморфозам, ни катаморфозам, поскольку общий уровень организации при этом существенно не меняется.
Такие эволюционные преобразования, которые значительно увеличивают адаптивные возможности данной группы (позволяя расширить старую или занять новую адаптивную зону), оказывают существенное влияние на организм в целом, но не изменяют общий уровень организации, были названы эпектоморфозами. Обеспечивая возможность существования в обширной адаптивной зоне, эпектоморфозы сохраняются длительное время в ходе последующей адаптивной радиации, становясь признаками крупных таксонов. Соответствующее направление эволюционных изменений было названо эпектогенезом. Соотношения между разными направлениями эволюционного процесса можно проиллюстрировать схемой (рис. 107), в основе которой лежит аналогичная схема А.Н.Северцова, модифицированная в связи с введением эпектогенеза.
Необходимо отметить, что все указанные формы эволюционных преобразований организмов (аро-, эпекто-, алло- и катаморфозы) определяются как таковые лишь в масштабе макрофилогенеза, когда приспособительный характер эволюционных изменений и их значение для организма как целого становятся вполне очевидными. На уровне же микроэволюции происходят как приспособительные (направляемые отбором), так и неприспособительные (нейтральные) изменения генофондов популяций; их дальнейшая эволюционная судьба и значение для организма, как правило, еще недостаточно ясны. Поэтому целесообразнее вообще не употреблять указанные термины для обозначения микроэволюционных изменений и использовать их лишь на макроэволюционном уровне.
Направляющие факторы эволюции
С точки зрения сторонников "оппортунистической эволюции", за различными направлениями эволюционных изменений не стоит никаких закономерных и организующих тенденций, кроме действия естественного отбора, который организует изменчивость организмов лишь в направлении развития приспособлений к изменениям среды обитания. С этих позиций главные направления эволюционного процесса (аро-, эпекто-, алло- и катагенез), по сути дела, равнозначны - в том смысле, что каждое из них является лишь средством для достижения успеха данной группы организмов в борьбе за существование (такой точки зрения придерживался и А.Н.Северцов).
Действительно, среди движущих факторов эволюции лишь естественный отбор обладает организующим воздействием на изменчивость организмов, и при этом отбор действительно лишен определенной направленности, что подчеркивал еще Ч.Дарвин. Но Дарвин указал и на фактор, определяющий конкретные направления эволюционных преобразований: "Природа условий имеет в определении каждого данного изменения подчиненное значение по сравнению с природой самого организма". Хотя эволюция организмов основывается на вероятностных процессах - возникновении мутаций (фенотипическое проявление которых неадекватно вызвавшим их появление изменениям внешних условий) и естественном отборе, "природа организма", т. е. организационная основа живых систем, ограничивает проявления случайности в эволюции определенными рамками. Иначе говоря, системная организация канализует филогенез, т.е. направляет эволюционные преобразования в определенные русла, и для любой конкретной группы организмов выбор возможных путей эволюции ограничен. Концепция жестко запрограммированной (номогенетической) эволюции основана на абсолютизации направляющей роли в эволюционном процессе организационной основы живых систем, тогда как концепция оппортунистической эволюции - на абсолютизации направляющей роли естественного отбора. Обычно истину следует искать где-то между крайними точками зрения.
Конкретные направления филогенетических преобразований различных групп организмов определяются взаимодействием сил естественного отбора и исторически сложившейся организации этих групп. Поэтому можно говорить о двух категориях направляющих факторов эволюции: внеорганизменных (силы отбора) и организменных.
Для любого данного вида особенности его организации создают предпосылки (преадаптации) к развитию определенных приспособлений и препятствуют развитию других, "разрешают" некоторые направления эволюционных преобразований и "запрещают" другие направления. Совокупность этих позитивных и негативных характеристик эволюционных возможностей данной группы и обозначена как организменные направляющие факторы эволюции. Эти факторы можно разделить (несколько условно) на три категории, в соответствии с уровнем их проявления в онтогенезе: 1) генетические, 2) морфогенетические, 3) морфофизио-логические (морфофункциональные).
Действие двух первых категорий организменных направляющих факторов в полной мере проявляется уже на уровне микроэволюции. Как уже отмечалось (ч. II, гл. 1), каждый генотип и генофонд каждого вида характеризуется определенным набором возможных ("разрешенных") мутаций, или спектром мутационной изменчивости, который ограничен не только качественно, но и количественно, т. е. определенной частотой возникновения каждого типа мутаций. При этом некоторые мутации оказываются для данного генотипа (и генофонда) невозможными (запрещенными) - например, синие и зеленые окраски глаз для мух-дрозофил или синие окраски цветов для розоцветных растений. Причиной этого является отсутствие в генотипе соответствующих биохимических предпосылок.
Так как генофонды родственных видов сохраняют наборы гомологичных генов, унаследованных от общего предка, в них закономерно появляются гомологичные мутации (см. с. 65). Гомологичные мутации могут послужить основой для параллельных эволюционных изменений у близкородственных видов, относительно недавно дивергировавших от общего предка. Однако с течением времени в генофондах обособленных видов неизбежно накапливаются разнокачественные (негомологичные) мутации; это происходит даже при действии стабилизирующего отбора, когда фенотипический эффект мутаций структурных генов блокируется генами-модификаторами. У разных видов, генофонды которых были изолированы друг от друга достаточно длительное время, сохраняются гомологичные структуры фенотипа, но их генетический контроль может значительно (и даже практически полностью) различаться. Поэтому параллельная эволюция филетических линий, давно дивергировавших от общего предка (до уровня разных родов, семейств и т.д.), основывается не столько на гомологичных мутациях, сколько на действии двух других категорий организменных направляющих факторов.
(Гомология - сходство структур, основанное на общности их происхождения. Соотношение гомологичных структур, принадлежащих к разным уровням иерархической организации биологических систем (в том числе генетическая и фенотипическая гомология), имеет сложный и неоднозначный характер).
Некоторые мутации, возможные для данного генотипа в биохимическом отношении (т. е. разрешенные на генетическом уровне), в конечном итоге, однако, приводят к гибельным для развивающегося организма последствиям в виде нарушений морфогенеза (летальные мутации, например, морфогенетические последствия мутации врожденной гидроцефалии у домовой мыши, см. с. 325). Каждый онтогенез может быть изменен лишь определенным образом, т. е. в рамках соответствующего спектра возможных онтогенетических изменений. Это еще более сужает выбор возможных направлений эволюционных преобразований.
Наконец, существуют также морфофизиологические эволюционные ограничения и запреты, действие которых (так же как и соответствующих преадаптации) в полной мере проявляется лишь в масштабах макроэволюции, являясь одной из специфических причин ее направленного характера. Они обусловлены разнообразными взаимосвязями в пределах морфофизиологических систем и между этими системами в фенотипе взрослых организмов. При этом мутации и перестройки онтогенеза, которые могли бы привести к соответствующим изменениям фенотипа, сами по себе вполне возможны, и мутантные особи с определенной частотой могут появляться в популяциях данного вида. Однако возникающие при этом изменения фенотипа (даже, казалось бы, обладающие высокой приспособительной ценностью!) не могут быть использованы для формирования новых адаптации из-за своего несоответствия морфофизиологической организации данного вида. Такие преобразования остаются неосуществимыми до снятия соответствующих морфофизиологических запретов.
Так, например, в эпидермисе земноводных могут развиваться ороговения -- имеются необходимые для этого биохимические предпосылки, отсутствуют и морфогенетические запреты для этого процесса. И действительно, локальные ороговения эпидермиса развиваются в покровах некоторых видов амфибий (например, роговые когти у шпорцевых лягушек или у самцов когтистых тритонов, роговые "зубы" у головастиков многих видов бесхвостых амфибий). Однако для земноводных оказалось невозможным сформировать на этой основе такие ороговения покровов, которые могли бы эффективно защищать организм от обезвоживания в воздушной среде и в соленых водоемах, как у рептилий, птиц и млекопитающих. Это обусловлено необходимостью для земноводных сохранять постоянно влажную поверхность кожи, которая используется как дополнительный орган газообмена, в первую очередь для выведения из организма углекислого газа (подробнее см. ниже).
Морфофизиологические эволюционные ограничения и запреты обусловлены необходимостью гармонических перестроек систем организма, интегрированных адаптивно (т.е. входящих в общий адаптивный комплекс), функционально или хотя бы топографически. В филогенезе действие таких ограничений проявляется в форме различных координаций (т.е. филогенетических корреляций) между различными структурами и системами организма. Под топографическими координациями понимаются наиболее простые сопряженные эволюционные изменения органов, тесно связанных пространственно. Например, увеличение размеров глаз невозможно без соответствующих перестроек черепа, изменений положения мышц, сосудов и нервов в глазнице и височной области. Динамические координации представляют собой филогенетические взаимоотношения органов, связанных друг с другом в онтогенезе функциональными корреляциями. Примером эволюционных ограничений, основанных на таких координациях, является невозможность усиления какой-либо группы мышц без соответствующего усиления структур скелета и некоторых других мышечных групп, поскольку это сделало бы согласованную работу скелетно-мышечной системы механически несовершенной. Так, нет смысла в развитии мощных бедренных мышц при сохранении более слабых мышц голени, так как последние не могут эффективно передавать на субстрат силу сокращения первых. В то же время мышцы голени не могут значительно усиливаться у животных, приспособленных к быстрому бегу, поскольку это существенно увеличило бы момент инерции конечности. Это эволюционное ограничение требует развития у быстро бегающих животных характерной конструкции конечностей, в которых основная масса мышц располагается в проксимальных отделах (плечо, бедро), а сила их сокращения передается к опоре через тонкие и легкие дистальные отделы (предплечье, голень, стопа) посредством системы сухожилий.
И.И.Шмальгаузен выделял еще так называемые биологические координации, под которыми понимаются сопряженные изменения органов и отдельных структур, не связанных друг с другом непосредственно какими-либо корреляциями в онтогенезе, но входящими в общий адаптивный комплекс (например, эволюционные взаимосвязи между строением жевательных мышц, зубов, челюстных костей и челюстных суставов, обусловленные определенным способом питания). Координированные эволюционные изменения этих гетерогенных структур определяются естественным отбором.
Ключевые признаки и каскадные взаимодействия
В целостном организме различные органы и структуры взаимосвязаны различными формами корреляций и координации, и цепи этих взаимосвязей сложно переплетаются друг с другом. При этом в организации многих групп организмов возникают своего рода узкие места - такие ключевые морфофункциональные особенности организации, которые через каскад различных коррелятивных и координационных взаимосвязей оказывают определяющее влияние на особенности функционирования многих зависимых систем организма.
В координационном каскаде взаимосвязанных систем состояние каждого последующего уровня определяется предшествующим, начиная от ключевой, или лимитирующей, системы. Преобразования ключевой системы могут оказывать как негативное ("запрещающее"), так и позитивное ("разрешающее") воздействие на эволюционные изменения зависимых систем организма. В первом случае они обусловливают возникновение морфофизиологических эволюционных запретов, препятствующих развитию определенных адаптации и направлений эволюционных преобразований, во втором, наоборот, снимают подобные запреты и ограничения, существовавшие прежде.
Очень интересны каскадные морфофизиологические взаимосвязи в организации земноводных, определяемые особенностями их дыхательной системы. Земноводные используют для вентиляции легких нагнетательный насос, образованный подъязычным аппаратом и его мускулатурой и представляющий собой модифицированный жаберный насос предков наземных позвоночных - кистеперых рыб. Несовершенство этого дыхательного насоса, расположенного впереди легких (при отсутствии эффективных механизмов, способных изменять объем самих легочных мешков), приводит к неполноте опорожнения легких при выдохе, к сохранению там некоторого количества застойного воздуха, к смешению отработанного и свежего воздуха в ротоглоточной полости (рис. 108). В результате хотя легкие у земноводных могут обеспечить организм достаточным количеством кислорода, но они неспособны эффективно освобождать его от углекислого газа. Это сделало необходимым развитие у земноводных дополнительного пути выведения углекислого газа - через кожу. Как мы уже упоминали, дыхательная функция кожи требует постоянной влажности покровов за счет деятельности кожных желез и запрещает интенсивное ороговение эпидермиса. Это резко ограничивает круг доступных для земноводных местообитаний как на суше (где амфибии активны, как правило, лишь при повышенной влажности), так и в воде: из-за высокой проницаемости покровов земноводные не могут осуществлять эффективную осморегуляцию в гипертонической среде соленых водоемов (где их организм обезвоживается, теряя влагу через покровы), и море для земноводных недоступно. Напротив, в гипотонической среде пресных водоемов через покровы в организм земноводных непрерывно поступает избыточное количество воды, которую должны удалять из организма органы выделения. Это препятствует развитию приспособлений для экономии воды в выделительной системе амфибий. Кровеносная система земноводных должна обслуживать два органа газообмена - легкие и кожу, причем кровь от них поступает к сердцу по разным магистральным сосудам (легочная вена, впадающая в левое предсердие, и кожные вены, несущие кровь через систему полых вен в правое предсердие). Это определяет необходимость смешивания обоих потоков крови в желудочке сердца и невозможность эффективного разделения артериальной и венозной крови. В результате этого для земноводных невозможно развитие эффективных систем терморегуляции и высокого уровня метаболизма, достижение гомойотермии для земноводных запрещено.

Очевидно, эволюцию дыхательной системы земноводных можно интерпретировать как пример инадаптивного варианта эволюционных преобразований, по В.О.Ковалевскому. В организации земноводных несовершенный способ вентиляции легких оказался ключевым эволюционным запретом для развития важных адаптации во многих зависимых системах организма.
Но нередко и весьма совершенное само по себе приспособление может сыграть роль ключевого запрета по отношению к развитию определенных адаптации других систем организма. Эффектный пример такого рода представляют каскадные взаимосвязи в организации насекомых, также определяемые особенностями их дыхательной системы. Органы дыхания насекомых образованы сложной системой трахей - разветвленных воздухоносных трубочек, пронизывающих все тело; тончайшие трахейные стволики достигают практически всех клеток. По мнению М.С.Гилярова, именно совершенство трахейной системы позволило насекомым освоить широкий круг наземных местообитаний в условиях дефицита влаги. Вместе с тем трахейная система создала предпосылки для значительной интенсификации газообмена, вплоть до достижения у некоторых видов насекомых временной гомойотермии при полете.
С другой стороны, как было показано В.Н.Беклемишевым, трахейная система определенным образом ограничивает адаптивные и эволюционные возможности насекомых, накладывая отпечаток на всю их организацию. Крайне разветвленные трахеи делают ненужным участие кровеносной системы в газообмене, и ее периферические отделы в значительной степени редуцируются. Но при таком устройстве дыхательной и транспортной систем затруднено снабжение массивных органов, что ограничивает возможности увеличения размеров тела. Огромное большинство насекомых имеет мелкие размеры (у крупных современных видов жуков, палочников, стрекоз длина тела не превышает 13-15 см, лишь у некоторых тропических палочников она достигает 26 см). "Эволюционные эксперименты" насекомых в области крупных форм были неудачны: самые крупные известные насекомые, стрекозы-меганевры, достигавшие около 70 см в размахе крыльев при длине тела около 30 см, вымерли еще в раннепермское время.
Таким образом, у насекомых специфические особенности распределительной и дыхательной систем стали причиной возникновения эволюционного запрета на увеличение размеров тела. Небольшие размеры тела, в свою очередь, определяют многие особенности организации, поведения и экологии насекомых, которые достигли высокого совершенства в "мире малых форм". Малые размеры тела ограничивают количество клеток в организме, в частности в центральной нервной системе. Для насекомых характерна автономизация периферических отделов нервной системы (замыкание рефлекторных дуг на уровне периферических ганглиев) и преобладание наследственно закрепленных автоматизированных форм поведения. Трахейная система ограничивает также возможности экологической экспансии насекомых: при огромном разнообразии форм и адаптации насекомые совершенно не смогли освоить наиболее древнюю и обширную среду жизни - толщу морской воды. Вероятно, это связано с высокой плавучестью тела насекомых, пронизанного густой сетью воздухоносных ходов.
Причины параллельной эволюции
Очевидно, виды, происходящие от общего предка и сохраняющие гомологичные гены, морфогенетические системы и структуры фенотипа, должны обладать по этим причинам значительным сходством направляющих эволюцию организменных факторов. Закономерным результатом этого является широкое распространение явлений параллельной эволюции родственных филетических линий, известное как одно из проявлений направленности филогенеза. При этом, как мы уже отмечали, гомология генов имеет существенное значение только для близкородственных видов, тогда как параллельная эволюция таксонов более высокого ранга определяется в первую очередь сходством морфогенетических и морфофизиологических факторов.
Длительной параллельной эволюции независимых филогенетических стволов в сильной степени способствуют унаследованные от общих предков каскадные взаимодействия между системами органов. Изменения ключевой системы сказываются на состоянии всего каскада зависимых от нее систем, создавая предпосылки для определенных направлений эволюционных преобразований и запрещая другие. После формирования какого-либо ключевого приспособления начинается адаптивная радиация. Возникшие при этом родственные филетические линии наследуют вместе с ключевым признаком весь комплекс обусловленных им системных взаимосвязей и соответственно весь набор эволюционных потенций, т.е. возможных направлений эволюционных изменений и эволюционных запретов. Неизбежным следствием этого является возникновение параллельных эволюционных преобразований в зависимых от ключевого признака системах организма у родственных филетических линий.
Таким образом, исторически сложившаяся организация данного таксона ограничивает для составляющих его видов выбор возможных направлений эволюции определенными рамками. Если естественный отбор длительное время благоприятствует развитию какой-либо категории адаптации (например, переработке пищи челюстями у хищников, питающихся крупной добычей, как у предков млекопитающих), направленность эволюционных преобразований в пучке родственных филетических линий получает, так сказать, "двойное обеспечение" - со стороны как организменных, так и внеорганизменных факторов. Это является причиной возникновения устойчивых эволюционных тенденций и параллельной эволюции независимых линий в филогенезе многих групп организмов.
Одним из важнейших проявлений направленности макроэволюции является поступательный характер и широкое распространение эволюционных преобразований по пути морфофизиологи-ческого прогресса. Эволюционный прогресс и связанные с ним проблемы мы рассмотрим в следующей главе.
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





