|
|
Отцовские хромосомы темные, материнские — светлые.Стр 1 из 4Следующая ⇒ А — аутосомы; X, Y — половые хромосомы.
В сперматогенезе принято различать четыре стадии: размножения, роста, созревания и формирования (рис. 13). Полный цикл развития половая клетка проходит у быка за 63 дня, у барана— за 48, у хряка — за 40 дней. Процесс сперматогенеза протекает волнообразно. На протяжении одного канальца можно последовательно проследить все его этапы (рис. 14). В пределах же среза в стенке канальца невозможно увидеть одновременно все виды клеток. В стадии размножения происходит многократное митотическое деление сперматогоний. При этом часть половых клеток сохраняет все черты эмбриональности, остается стволовыми клетками и в дальнейшем является источником пополнения сперматогенного эпителия. Другая часть вступает на путь дифференцировки. И те и другие клетки называются сперматогониями, хотя и различаются по морфологическим признакам. Все сперматогоний прилежат к оболочке извитого канальца, располагаясь на базальной мембране. Это уплощенные или овальные клетки со светлой цитоплазмой и округлыми или овальными ядрами. Хроматин, вначале мелкодисперсный, с каждым делением становится все заметнее. У животных каждого вида наблюдается определенное количество сперматогониальных делений (от 1 до 14), чаще 4—6 (у быка — 6, у барана — 5).
Отличительная особенность сперматогоний, вступивших на путь дифференцировки, — неполная цитотомия, в результате чего сестринские и дочерние сперматогоний оказываются связанными между собой цитоплазматическими мостиками, то есть являются синцитием (рис. 15). Мостики (фузомы) достаточно длинные, так что клетки могут отходить друг от друга на расстояние до 25 мкм. Все связанные между собой клетки образуют клон. Клетки одного клона синхронно проходят стадии сперматогенеза. После завершения сперматогониальных делений все клетки клона вступают в стадию роста. В стадии роста половая клетка называется сперматоцит I порядка. В это время в клетке идут активные процессы ассимиляции, дифференцировки органелл, синтез ДНК, размножение митохондрий делением, увеличивается ядрышко. Клетка округляется, оттесняется от базальной мембраны по направлению к просвету канальца, увеличивается в размерах. По количеству хромосом сперматоцит I порядка считается диплоидным, а по содержанию ДНК — тетраплоидным, так как каждая хромосома состоит из двух хроматид. В крупном округлом ядре его последовательно происходят преобразования хромосом, характерные для профазы I мейоза. Потеряв связь с базальной мембраной семенного извитого канальца, сперматоциты I порядка вступают в тесный контакт с поддерживающими клетками, которые называют также клетками Сертоли, фолликулярными, питающими, выстилающими, защитными и т. д. Это однослойный эпителий из очень крупных клеток, высотой во всю толщину стенки канальца с бледной цитоплазмой и ядрами неправильной формы, лежащими у базального полюса, с большим количеством ветвистых отростков. Поддерживающие клетки выполняют разнообразные функции (опорную, трофическую, защитную, гормональную, разграничительную, фагоцитарную, регуляторную, секреторную и др.) и строение их подвержено циклическим изменениям в зависимости от стадии развития половых клеток.
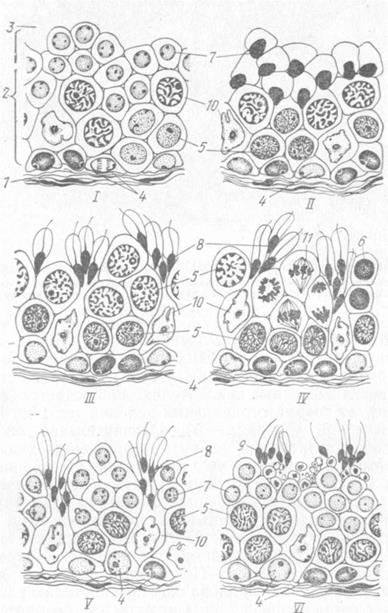
Рис. 15. Взаимоотношения между поддерживающими и половыми клетками: 1 — сперматогоний; 2 — сперматоцит I порядка; 3 — сперматида поздняя; 4 — мостик; 5 — поддерживающая клетка (сертоли); 6 — ее ядро; 7 —плотный контакт между двумя клетками сертоли.
В стадии созревания происходит редукция числа хромосом в половых клетках в процессе двух делений созревания, быстро следующих друг за другом, то есть в процессе мейоза. В результате первого деления созревания — редукционного деления мейоза из каждого сперматоцита I порядка образуется два сперматоцита II порядка. Это короткоживущие клетки, более мелкие, чем сперматоциты I порядка. Они содержат гаплоидное число хромосом (но каждая хромосома состоит из двух хроматид) и тут же вступают во второе деление созревания — эквационное деление мейоза. Между делениями созревания интерфаза очень короткая. В ней отсутствует S-период, нет деконденсации ДНК. А так как хромосомы сохраняют бивалентную структуру, профаза II проходит очень быстро. В результате второго деления из каждого сперматоцита II порядка образуются две сперматиды — истинно гаплоидные клетки. У них в сравнении с соматическими клетками уменьшено вдвое не только количество хромосом, которое гаплоидно (1n), но и количество ДНК (1с). Кроме того, сперматиды различаются по половым хромосомам: половина их содержит Х-хромосому, а половина Y-хромосому. Остальные хромосомы их отличаются и от родительских хромосом и друг от друга в разных клетках, так как произошел случайный обмен генами во время кроссинговера. Сперматоциты II порядка, и особенно сперматиды, находятся в тесном контакте с клетками Сертоли. В процессе созревания они перемещаются все ближе к просвету канальца и все глубже проникают в цитоплазму поддерживающей клетки. 1. Характеристика спермиев млекопитающих
В стадии формирования сперматиды превращаются в спермий (сперматозоид). Данную стадию проходят одновременно все сперматиды одного клона. На нее приходится около 1/3 времени всего сперматогенеза. Процессы, происходящие в половых клетках в стадии формирования, подробно изучены у некоторых видов млекопитающих (в основном у лабораторных животных). При этом выделено 19 этапов, которые проходит сперматида, прежде чем превратится в спермий. Сперматиды, только что появившиеся в результате второго деления созревания, — ранние сперматиды—это мелкие округлые клетки с небольшим округлым ядром, содержащим мелкогранулярный хроматин. В их цитоплазме имеются все органеллы общего значения. Сперматиды в период формирования располагаются в карманах — углублениях на апикальном полюсе клетки Сертоли, которые, вероятно, вырабатывают гормон, регулирующий их дифференцировку. В процессе дифференцировки на переднем полюсе сперматиды в области комплекса Гольджи развивается акросома — специфическая органелла, обеспечивающая растворение оболочек яйцеклетки благодаря содержанию большого количества гиалуронидазы и других гидролитических ферментов. Акросома примыкает к ядру и по мере развития прикрывает его. Ядро вытягивается, уплощается и резко уплотняется. Объем его уменьшается в 20—50 раз. Происходит конденсация и специальная упаковка хромосом с помощью белков — протаминов. Упаковка эта настолько плотная, что дезоксирибонуклеопротеиды приобретают даже некоторые свойства кристаллов. От центриоли отрастает жгутик, направленный в сторону, противоположную акросоме — к просвету канальца. Клетка принимает грушевидную форму. Часть протоплазмы сползает со сперматиды и фагоцитируется клеткой Сертоли. Сперматида все более удлиняется. Такая клетка называется поздней сперматидой. Когда процесс дифференцировки сперматиды завершается, она теряет связь с клеткой Сертоли и другими клетками своего клона и становится спермием.
Головка имеет видоспецифическую форму. У сельскохозяйственных животных она лепешкообразная. Акросома в виде шапочки покрывает головку на 2/3. Основу головки составляет ядро, в котором содержатся ДНК в виде плотно упакованных хромосом. Цитоплазма очень тонким слоем покрывает ядро. Шейка — самый короткий отдел, видимый только в электронный микроскоп. В ней располагается центросома из двух центриолей. Проксимальная центриоль — та, что ближе к ядру, лежит поперек шейки, а дистальная — более удаленная от ядра — вдоль шейки. От нее берет начало осевая нить (аксонема) жгутика. Хвост составляет основную длину клетки. Он состоит из среднего (промежуточного), главного и концевого отделов. На протяжении всего хвоста проходит осевая нить. Она подобно ресничкам состоит из двух центральных и девяти пар периферических микротрубочек. Микротрубочки построены из белков тубулина и динеина, благодаря которым осуществляется волнообразное и бичевидное движение жгутика. В среднем и главном отделах хвоста микротрубочки сопровождаются расположенными снаружи девятью дополнительными плотными фибриллами. В среднем отделе хвоста поверх фибрилл лежат митохондрии, упакованные в виде спирали. Это так называемая спиральная нить спермия. Средний отдел от главного отделяется замыкающим кольцом. Цитоплазма тонким слоем покрывает весь хвост, за исключением концевого отдела (последних 5—7 мкм). Иногда цитоплазма образует 1—2 гребневидных выроста — ундулирующую мембрану, дополнительное приспособление для передвижения. Созревание спермия. Сформированные спермин млекопитающих, находящиеся в семенниках, еще не могут выполнять специфическую функцию — они не обладают способностью к активному движению и оплодотворению. Из семенного извитого канальца спермин выносятся с током жидкости, заполняющей его просвет. Процесс созревания спермия продолжается во время его перемещения в половых путях самца и самки, вплоть до момента оплодотворения. При этом меняется строение оболочки спермия, его физико-химическое состояние. Так, пройдя придаток семенника спермий приобретают способность к активному движению, их оболочка покрывается углеводно-белковым слоем. В половых путях самки в спермиях происходят изменения, способствующие контакту спермия с яйцеклеткой, акросомной реакции и оплодотворению. При этом жизнеспособность спермиев в половых путях самки первоначально даже повышается (у спермиев быка в течение двух суток)—увеличивается потребление кислорода, вырабатывается больше энергии. Однако в дальнейшем жизнеспособность, оплодотворяющая способность спермиев резко падает — происходит их старение. У различных животных оптимальный срок пребывания спермиев в половых путях самок разный. У млекопитающих — это от одного до нескольких дней, у птиц — от одной до нескольких недель, у матки медоносной пчелы — до 3—5 лет. При оплодотворении «старыми» спермиями увеличивается процент эмбриональной смертности, уродств, снижается оплодотворяемость. Считают, что это вызвано излишней прочностью связи ДНК с белками в хромосомах у «старых» спермиев. Подобное следует учитывать при осеменении животных, синхронизируя время охоты и покрытия самок. Влияние различных факторов на сперматогенез. Продолжительность сперматогенеза постоянная для данного вида животных и не зависит от действия эндогенных и экзогенных факторов. У быка она равна 62—63 дня, у барана — 47—48 дней, у хряка — 39—40 дней. Что же касается активности сперматогенеза, качества и количества развивающихся половых клеток, то здесь прослеживается заметное влияние различных факторов. В процессе сперматогенеза из одной сперматогоний, вступающей на путь дифференцировки, теоретически может образоваться до 36 768Х215 спермиев. Однако этого не происходит, так как в сперматогенезе отмечено несколько волн гибели клеток, которые приходятся на стадии размножения (во время профазы митоза в сперматогониях), созревания (во время деления созревания) и формирования (во время конденсации хроматина в сперматидах). В результате спермиев образуется 50—70% от теоретически рассчитываемого количества. Причиной дегенерации клеток в период сперматогенеза считаются спонтанные нарушения хромосом в процессе деления. Высказывается также мнение, что на гибель клеток влияет нарушение соотношения между половыми и поддерживающими клетками. Различные внешние воздействия: жесткое ультрафиолетовое, рентгеновское облучение и другие виды проникающей радиации, низкая или высокая температуры, обездвиживание, удаление гипофиза, гипоксия, введение гормонов (эстрогенов) и других биологически активных веществ, общие тяжелые заболевания, травма и пересадка семенника, крипторхизм приводят к однотипной реакции. Все эти и многие другие факторы вызывают или тотальную гибель сперматогенного эпителия, или клеток на определенной стадии развития и тем угнетают сперматогенез; увеличивают число атипичных клеток, неспособных к оплодотворению (с измененным ядром, центросомой, с измененным содержанием ДНК, с нарушениями веретена деления, митоза и мейоза). Наиболее чувствительны клетки в период конъюгации хромосом (профаза I мейоза) и во время второго деления созревания. С. С. Райцина на основании многочисленных исследований приходит к выводу, что однотипная реакция сперматогенного эпителия на повреждающие внешние воздействия вызвана нарушением гематотестикулярного (haema — кровь, testis — семенник) барьера, регулирующего поступление из крови в семенные канальцы сывороточных белков, биологически активных веществ, метаболитов, токсинов, лекарственных препаратов. Барьерную функцию в семеннике выполняют: белочная оболочка, собственная оболочка семенных извитых канальцев и клетки Сертоли. Нарушение гематотестикулярного барьера приводит к гибели или атипичному развитию сперматогенного эпителия. Гематотестикулярный барьер у новорожденных животных несовершенен и достигает полного развития к началу пубертатного периода, что следует, учитывать при выращивании самцов-производителей. В консервирующих смесях спермин могут долго сохранять жизнеспособность. Данное качество используется при искусственном осеменении животных. Есть опыт хранения спермы быка до 25 лет, барана — до 11, хряка — до 3 лет. Это имеет большое значение для развития животноводства, а также для сохранения генофонда редких и исчезающих видов.
Развитие женских половых клеток — оогенез
Женская половая клетка — яйцеклетка (ovum — яйцо) — была открыта К. Бэром в 1825 г. Развитие женских половых клеток— оогенез, происходит в яичнике и завершается в яйцеводе. В оогенезе различают три стадии: размножения, роста и созревания (см. рис. 13). Стадию размножения женских половых клеток у млекопитающих отмечают в утробный период жизни особи. Первичные половые клетки, вселившись в закладку яичника, превращаются в оогонии и много раз подряд делятся митозом — идет накопление половых клеток. Оогонии — круглые или овальные клетки с оксифильной цитоплазмой и крупным ядром, в котором видны 1—2 ядрышка и мелкодисперсный хроматин. Среди соматических клеток яичника они выделяются своими крупными размерами.
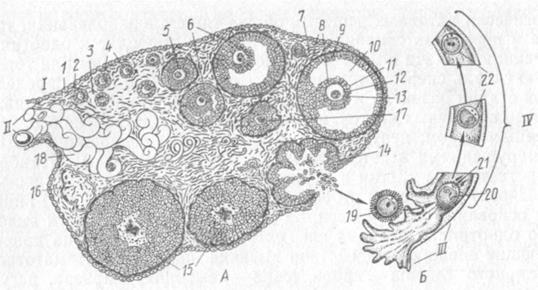
Рис. 17. Рост и созревание женских половых клеток в яичнике (А) и яйцеводе (Б): I — корковое вещество яичника; II — мозговое вещество; III — воронка и IV — просвет яйцевода; 1 — покровный эпителий; 2 — белочная оболочка; 3 — интерстициальная ткань; 4 — первичный (примордиальный) фолликул; 5 —растущий (вторичный) фолликул; 6 — пузырчатый фолликул; 7 — тека фолликула; 8 — фолликулярный эпителий; 9 —зернистый слой; 10 — полость фолликула; 11 — лучистый венец; 12 — блестящая оболочка; 13 — ооцит I порядка; 14 — овулированный фолликул; 15 — желтое тело; 16 — беловатое тело; 17 — атретический фолликул; 18 — сосуды; 19 — ооцит II порядка; 20 — зрелая яйцеклетка; 21 —полярное тельце; 22 — зигота. К концу плодного периода у млекопитающих большинства видов митотическая активность оогониев затухает. У кроликов и свиней, по данным некоторых исследователей, размножение оогониев продолжается некоторое время после рождения (у свиней — в течение первого месяца). Прекратив делиться, половые клетки вступают в стадию роста (рис. 17). Стадия роста оогенеза проходит в яичнике, отличается длительностью и сложностью преобразований. Половая клетка на этой стадии называется ооцитом I порядка. В его ядре происходят преобразования, характерные для профазы I мейоза. У большей части ооцитов I порядка отмечают синхронные последовательные стадии профазы I, вплоть до диплонемы. При этом половая клетка окружается одним слоем клеток — фолликулярным эпителием. Образуется многоклеточная структура — фолликул. Фолликул, стенки которого образованы одним слоем плоских фолликулярных клеток, плотно прилегающих к ооциту I порядка, называется примордиальным, а затем, когда фолликулярные клетки увеличиваются в размерах, — первичным. В первые дни после рождения ооциты проходят диплонему и вступают в продолжительную интерфазную стадию диктиотены, предшествующую диакинезу. Она длится до полового созревания и заканчивается незадолго перед овуляцией. В это время хромосомы становятся невидимыми, их ДНК деконденсируется, активно идет синтез всех видов РНК, различных белков, увеличивается количество органелл, большинство которых располагается вокруг ядра. В результате всех этих процессов происходит рост ооцита I порядка. Рост совершается благодаря ассимиляционной деятельности половой клетки. Идет он медленно, продолжается долго— месяцы и годы и называется периодом малого, или цито-плазматического роста. Первичные фолликулы с ооцитами I порядка в стадии малого роста располагаются по периферии корковой зоны яичника. У взрослой коровы они составляют почти 55% от всех фолликулов яичника. Постепенно фолликулярные клетки увеличиваются в размерах и начинают активно функционировать. Такой фолликул называется растущим (вторичным). У коровы вторичные фолликулы составляют 43%. При совместной деятельности фолликулярных клеток и ооцита I порядка вокруг половой клетки формируется блестящая оболочка — вторичная оболочка половой клетки. Через ее поры устанавливается тесная связь половой и фолликулярных клеток. Последние, размножаясь, покрывают ооцит I порядка несколькими слоями. Особенно активен фолликулярный эпителий в период накопления желтка — в период вителлогенеза (vitellus — желток, genesis — происхождение), который начинается при половом созревании. С помощью фолликулярного эпителия идет быстрое накопление ооцитом I порядка белковых и других веществ (вителлогенина, фосвитина, липовителлина, вителлина), синтезированных за пределами яичника, в основном в печени и принесенных кровью. Период вителлогенеза называют еще периодом большого роста, или трофоплазматическим. У свиньи он протекает в фолликулах с диаметром от 4 до 8 мм. В это время происходит пропорциональное увеличение ооцита I порядка и фолликула. Фолликулярные клетки осуществляют отбор и концентрацию вокруг ооцита молекул, подлежащих эндоцитозу. Они же, по-видимому, выделяют вещества, стимулирующие микропиноцитоз в ооците и гормоны, стимулирующие синтез вителлогенина печенью. Живые неокрашенные ооциты I порядка в период большого роста имеют коричневый цвет. Органеллы в них располагаются в основном по периферии цитоплазмы, где в первую очередь идет отложение желточных гранул и других включений. Постепенно желток заполняет большую часть цитоплазмы ооцита, вителлогенез затухает. Рост ооцита прекращается, а фолликул продолжает расти. Между фолликулярными клетками появляется полость, в которой накапливается жидкость, содержащая половые гормоны (эстрогены) и другие продукты деятельности фолликулярного эпителия. Фолликул становится пузырчатым (Граафов пузырек или третичный фолликул). У коровы полость начинает появляться в фолликулах, достигших диаметра 1 мм. Место, где в стенке фолликула расположен ооцит I порядка, называется яйценосным бугорком. Ооцит покрыт слоем фолликулярных клеток или лучистым венцом, который у сельскохозяйственных животных образован 8—12 рядами клеток. Остальные фолликулярные клетки формируют зернистый слой фолликула—гранулезу. Поверх него фолликул одет соединительнотканной оболочкой — текой. Тека выполняет опорную и трофическую функции, в ней много сосудов и нервов, залегают клетки, вырабатывающие половые гормоны. Созревший пузырчатый фолликул имеет макроскопические размеры (табл. 2).
  Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем...  Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? - задался я вопросом...  Живите по правилу: МАЛО ЛИ ЧТО НА СВЕТЕ СУЩЕСТВУЕТ? Я неслучайно подчеркиваю, что место в голове ограничено, а информации вокруг много, и что ваше право...  Что делать, если нет взаимности? А теперь спустимся с небес на землю. Приземлились? Продолжаем разговор... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|

 Строение спермия. Спермий — свободная, подвижная клетка, длиной у млекопитающих 40—60 мкм (табл. 1). Состоит из головки, шейки и хвоста. Все его части покрыты цитолеммой (рис. 16).
Строение спермия. Спермий — свободная, подвижная клетка, длиной у млекопитающих 40—60 мкм (табл. 1). Состоит из головки, шейки и хвоста. Все его части покрыты цитолеммой (рис. 16).