|
ГЛАВА 5. ТЕМПЫ МАКРОЭВОЛЮЦИИ
Среди специфических черт макроэволюции пульсирующий характер ее темпов, выражающийся в виде чередования периодов типогенеза и типостаза, более всего привлекал внимание исследователей. Именно эта закономерность макрофилогенеза способствовала возникновению сальтационных гипотез. Как уже отмечалось, типогенез имеет характер квантовой эволюции, при которой значительные преобразования организации в процессе формирования основного комплекса признаков нового крупного таксона происходят в течение относительно краткого промежутка времени. Напомним, что о краткости сроков здесь говорится в масштабах макрофилогенеза, для которого относительно краткими можно считать промежутки времени в 10-30 млн лет по сравнению с сотнями миллионов лет периодов типостаза. Выше (гл. 2 этой части) мы рассмотрели ряд гипотез, предложенных в рамках синтетической теории эволюции для объяснения высоких темпов типогенеза (смена адаптивных зон, "генетические революции", видовой отбор, биоценотическая регуляция и преобразования экосистем). Эти гипотезы.(особенно первая и последняя из них) вносят определенный вклад в решение данной проблемы, но каждая из них имеет отношение лишь к некоторым частным случаям.
Вероятно, квантовый эффект в макрофилогенезе является внешним выражением действия нескольких эволюционных механизмов, основанных на системных свойствах организмов и объединяемых пороговым характером своего проявления.
Эволюционные преобразования корреляционных систем
Самый простой вариант такого механизма представляют эволюционные перестройки функциональных (эргонтических) корреляционных систем фенотипа. В целостной корреляционной системе изменение одного компонента требует соответствующих изменений других компонентов. Например, невозможно изменить форму суставной поверхности на одной кости без соответствующих изменений другой кости, сочлененной с первой (см. также примеры динамических координации, с. 366). В сущности, коррелятивные связи в морфофункциональных системах организма сами являются причиной возникновения сил мощного направленного отбора, требующих гармонических перестроек целого по принципу "все или ничего".
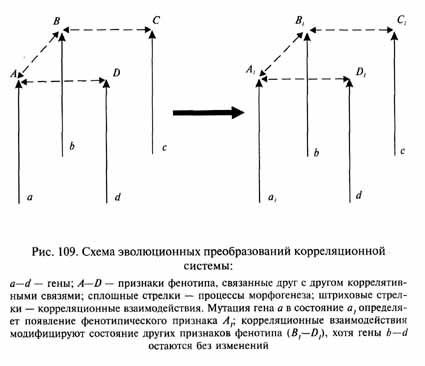
При этом очевидно, что одновременное или строго последовательное возникновение всех необходимых мутаций разных структурных генов, которые могли бы обеспечить гармоничное преобразование всех затронутых компонентов фенотипа, чрезвычайно маловероятно. Поэтому на первых этапах эволюционных преобразований таких корреляционных и координационных систем важную роль могут играть модификационные (ненаследственные) изменения взаимосвязанных элементов (рис. 109). Хотя такие изменения остаются в пределах прежней нормы реакции, они в совокупности могут привести к существенным преобразованиям общего фенотипического облика в очень короткие филогенетические сроки. Для палеонтолога, изучающего ископаемые остатки организмов, такие преобразования будут выглядеть как макроэволюционные.
Проиллюстрировать действие этого механизма проще всего на примере модификаций, компенсирующих различные повреждения организма, возникающие в естественных условиях или в эксперименте. Г. Понтер и Д.Уорд обнаружили среди отловленных в море окунеобразных рыб, принадлежащих к разным видам (Chaetodipterus faber, Micropogon undulatus и др.), несколько особей, у которых был полностью утрачен хвостовой плавник (вероятно, в результате нападения хищника), причем отсутствие хвоста было компенсировано разрастанием назад спинного и анального плавников и их мускулатуры. Эти рыбы были вполне жизнеспособны и, очевидно, прожили уже значительное время после утраты хвостового плавника. Конечно, в данном случае произошло не наследственное изменение организации, а лишь комплекс моди-фикационных ненаследственных изменений, компенсирующих повреждение. Однако можно полагать, что если утрата хвостового плавника будет обусловлена мутацией, то организм сможет компенсировать ее неблагоприятные последствия посредством аналогичных модификационных перестроек.
Интересные модификации скелетно-мышечной системы возникают у четвероногих млекопитающих в ответ на экспериментальное приучение к постоянному передвижению на задних конечностях. У обезьян-макаков это приводило к возникновению характерного для человека S-образного изгиба позвоночника; у крыс - к некоторому укорочению (брахицефализации) черепа, а также к усилению ягодичных мышц и изменениям формы таза, связанным с увеличением угла между лобковой и подвздошной костями. Особенно любопытно, что эти модификации в определенной степени сходны с изменениями соответствующих структур в эволюции предков человека, несомненно, связанными с переходом к двуногому передвижению.
В этом отношении интересны также результаты экспериментов И.Гармса с илистыми прыгунами (Periophthalmus) - окунеобразными рыбами (рис. ПО), обитающими в тропической приливо-отливной мангровой зоне. Во время отливов илистые прыгуны регулярно остаются на суше, где они могут переползать и даже совершать прыжки, отталкиваясь передними плавниками. Гармс выдерживал илистых прыгунов длительное время на суше и подвергал их воздействию повышенной концентрации тироксина (гормон щитовидной железы). В результате у рыбок уплотнялись ткани внутренних органов и кожа, усиливалось ороговение эпидермиса, удлинялись конечности, значительно уменьшались отолиты (слуховые камешки во внутреннем ухе, которые у рыб обычно имеют очень крупные размеры). Эти адаптивные модификации сходны с соответствующими изменениями, происходящими в онтогенезе земноводных при метаморфозе, который стимулируется повышенной секрецией тироксина щитовидной железой. Можно полагать, что эти изменения, приспосабливающие организм земноводных к жизни на суше, также могли первоначально представлять собой модификации, возникавшие в ответ на повышенную тироксинизацию, а эта последняя была результатом мутации, обусловившей усиление функции щитовидной железы.
Если возникшие подобным образом изменения фенотипа имеют адаптивную ценность, стабилизирующий отбор будет благоприятствовать наследственному закреплению новой комбинации фенотипических признаков и автономизации их онтогенетического развития. При этом отбор способствует закреплению и распространению мутаций, обеспечивающих формирование этих признаков в онтогенезе без модифицирующего влияния внешних условий - т. е. генохопий соответствующих модификаций. В связи с этим напомним подмеченный Р. Гольдшмидтом параллелизм фе-нотипического проявления мутаций и модификаций (см. с. 61). Дальнейшие перестройки онтогенеза в направлении автономизации развития этих признаков приведут ко все более ранней их закладке в эмбриогенезе. Отбор также будет способствовать новому расширению нормы реакции в том же направлении для создания нового "запаса прочности", нового резерва модификационной изменчивости, который может быть использован в последующей эволюции.
Таким образом, целостные системы организма, связанные функциональными корреляциями, преобразуются в эволюции как целое. Поэтому, как показал Л.Ридл, их преобразования должны происходить гораздо быстрее, чем при отборе независимых случайных вариаций отдельных признаков.
Каскадные корреляции и ключевые изменения
Важное значение для возникновения высоких темпов типоге-неза имеют каскадные взаимодействия между разными системами организма, рассмотренные в двух предшествовавших главах. Ключевые изменения являются своего рода "спусковым механизмом", запускающим весь каскад эволюционных преобразований в зависимых системах, имеющих важное адаптивное значение (как мы уже отмечали, эти преобразования были и раньше возможны на генетическом и морфогенетическом уровнях, но запрещены на морфофизиологическом, см. с. 365). После приобретения ключевого изменения при сильном давлении отбора, благоприятствующего развитию этих важных адаптации, эволюционные изменения должны происходить с максимально возможной скоростью. Таким образом, ключевые изменения организации являются своего рода "эволюционным триггером", значительно увеличивающим темпы макроэволюционных преобразований.
Происходящие при этом микроэволюционные изменения ничем не отличаются от обычного видообразования, и темпы микроэволюции в периоды каскадных макроэволюционных преобразований могут оставаться на среднем уровне. Однако взаимозависимость перестроек разных систем организма, скоординированных и разрешенных ключевым преобразованием, сама становится источником для возникновения мощного давления направленного отбора, благоприятствующего изменениям в заданном направлении. Скоординированность и направленность преобразований системной организации значительно ускоряют макроэволюцию.
Одним из наиболее известных примеров является каскад эволюционных преобразований у предков рептилий, связанный с развитием у них ключевого ароморфоза - механизма дыхательных движений грудной клетки, ставшего высокоэффективным всасывающим легочным насосом. Как мы видели в предыдущей главе, развитие этого насоса ликвидировало весь комплекс мор-фофизиологических ограничений и запретов, существующих в организации амфибий и обусловленных недостаточностью примитивного нагнетательного легочного насоса (подъязычного аппарата) для выведения из легких углекислого газа.
Возникновение направленности и в силу этого - высоких темпов макроэволюционных преобразований может быть обусловлено не только каскадными взаимодействиями, но и другими формами координации между различными системами организма - без ясно выраженного доминирования одной системы над другими (хотя квантовый эффект при этом, очевидно, будет выражен слабее). Такой характер макроэволюции соответствует обоснованной Т. Кемпом модели взаимосвязанного прогресса, согласно которой формирование организации нового крупного таксона происходит путем серий взаимосвязанных изменений различных систем организма. По этой модели происходило развитие основных признаков организации млекопитающих в филогенезе высших синапсидных рептилий — териодонтов. При этом отсутствие общего межсистемного ключевого признака не исключает каскадных взаимодействий в отдельных системах.
Так, сложный комплекс преобразований челюстного аппарата у териодонтов (см. рис. 62) был обусловлен ключевой адаптацией - совершенствованием переработки пищи челюстями.
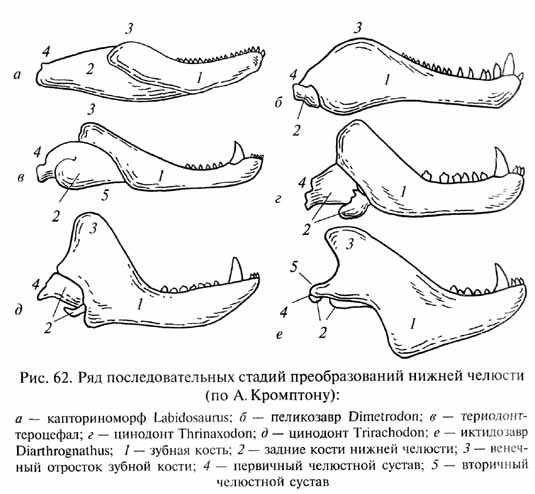
Эти преобразования включают дифференциацию зубов на резцы, клыки и трехбугорчатые щечные зубы ("коренные"), увеличение размеров зубной кости нижней челюсти и ее венечного отростка, на который прикреплялась височная мышца (роль последней усиливалась при развитии механизма пережевывания пищи). Развитие этих признаков непосредственно повлекло за собой изменение обших пропорций черепа (увеличение объема височных ям, вмещающих жевательную мускулатуру, привело к относительному увеличению заглазничной части черепа), развитие вторичного челюстного сустава, разрастание зубной кости и редукцию остальных костей нижней челюсти и преобразование костей первичного челюстного сустава в косточки среднего уха. Пережевывание пищи требует жесткого давления на перерабатываемый корм, что привело к утрате примитивной подвижности нёба и верхней челюсти (кинетизма черепа). С другой стороны, задержка пищи для переработки в ротовой полости приводит к задержке дыхания, крайне нежелательной при развитии гомойо-термии. Это вызвало необходимость развития вторичного нёба, разделившего носовой ход и собственно ротовую полость. Кроме того, вторичное нёбо укрепляет верхнюю челюсть, что также важно при пережевывании пищи. Развитие всех этих преобразований было обусловлено указанной ключевой адаптацией. Весь комплекс изменений развивался скоррелированно и направленно, что доказывается многочисленными параллелизмами в разных филетических линиях териодонтов. Это определило высокие темпы типогенеза.
Ускорению темпов типогенеза могут способствовать и другие эволюционные механизмы, обладающие пороговым характером проявления. Например, таковым является механизм морфофунк-циональной преадаптации, посредством которой в филогенезе происходит смена адаптивной роли органов. После прохождения стадии преадаптационного порога перед данной группой организмов открываются новые адаптивные возможности, которые реализуются в относительно краткие (в масштабах макроэволюции) сроки. Одновременно происходит перестройка всего комплекса структур, связанных корреляциями с органом, приобретшим новую адаптивную роль.
Возникновению квантового макроэволюционного эффекта могут также способствовать дискретные изменения программ онтогенеза. Взаимодействие различных факторов в морфогенезе часто имеет пороговый характер. В этом случае сравнительно незначительные генетические перестройки, обусловленные мутациями генов-регуляторов (особенно типа гомеобоксных генов, контролирующих экспрессию целых генетических систем) могут привести к существенным преобразованиям фенотипических признаков. Например, при обработке эмбриональных зачатков кожи ретиноевой кислотой (витамин А) зачатки роговых чешуи на конечностях у эмбрионов птиц преобразуются в перья, а зачатки волос у эмбрионов млекопитающих - в железистые структуры.
В эволюции корреляционных систем изменения их концевых звеньев и боковых ветвей могут происходить более или менее независимо друг от друга - вплоть до соответствия принципу независимой эволюции разных признаков по Г. Осборну. Скорость этих изменений определяется только факторами микроэволюции, в первую очередь частотой мутаций и давлением отбора. Такие изменения происходят наиболее часто и являются основой для процессов видообразования и адаптивной радиации, не затрагивающих основной план строения крупного таксона.
Гораздо реже возникают изменения на более глубоких уровнях организации корреляционных систем. Их следствием являются координированные изменения всех взаимосвязанных компонентов. В результате происходят дискретные преобразования целого комплекса признаков, в том числе в общем плане строения. Эти преобразования также осуществляются через обычные микроэволюционные механизмы, но их координированный и направленный характер обусловливает высокие темпы макроэво-люционных перестроек. Р. Ридл показал, что темпы эволюционных преобразований интегрированных целостных систем могут быть на несколько порядков выше, чем эволюция, основанная на комбинировании независимых вариаций отдельных компонентов.
Очевидно, при необходимости быстрых координированных перестроек гетерогенного адаптивного комплекса отбор должен благоприятствовать возникновению корреляционных связей между разными его компонентами и подчинению разных признаков общему контролю в онтогенезе, вплоть до формирования целостных генетических комплексов. При этом и на молекулярном уровне формируется блочно-иерархическая организация генома, позволяющая существенно увеличить скорость эволюции за счет действия отбора на целостные блоки в геноме, контролируемые общими генами - операторами и регуляторами (см. с. 90). Иначе говоря, длительно действующий направленный отбор может благоприятствовать более глубокой интеграции систем организма, которая способствует значительному убыстрению макроэволюции, поскольку эволюционные преобразования целостной системы происходят несравненно быстрее, чем комбинирование благоприятных мутаций по отдельным признакам.
Таким образом, можно заключить, что высокие темпы типогенеза определяются прежде всего самим характером происходящих изменений организации, причем осуществляется своего рода самоконтроль темпов макроэволюции со стороны организма.
Причины макроэволюционного типостаза
Все формы корреляционных и координационных связей в целостном организме при возникновении изменений в ключевых системах способствуют комплексному характеру макроэволюци-онных преобразований, что многократно повышает темпы типогенеза. С другой стороны, стабильность общего плана строения в период типостаза обеспечивается разными механизмами системного гомеостаза, действующими на всех уровнях организации, - канализация морфогенеза под воздействием генов-модификаторов, регуляция возникающих изменений при взаимодействиях разных морфогенетических систем, мультифункциональность и функциональная пластичность фенотипических структур и др. (Мультифункциональность - выполнение данным органом или структурой нескольких функций, связанных с разными адаптациями. В этом смысле термин "мультифункциональность" иногда заменяют на "мультиадаптивность").
Вообще степень эволюционного консерватизма данной системы в целом пропорциональна ее способности к авторегуляции. Например, в скелетно-мышечном комплексе мускулы обладают более высокой онтогенетической пластичностью, чем кости; в онтогенезе строение и функции мышц относительно легко модифицируются в соответствии с изменениями нагрузок. К тому же мышцы обладают высокой функциональной лабильностью, т.е. способны обеспечивать различные оттенки движений сокращением разных пучков мышечных волокон в различных комбинациях. Иными словами, мышцы обладают высокими авторегуляционными возможностями. Следствием этого является возможность для данного мышечного комплекса удовлетворительно функционировать при разнообразных адаптивных требованиях, соответствующих различным направлениям отбора. Скелетные же элементы обладают авторегуляционными способностями в значительно меньшей мере, чем мышцы. Поэтому они не могут адекватно реагировать адаптивными модификациями на существенные изменения условий функционирования и должны отвечать на изменения направления отбора наследственными перестройками, основанными на соответствующих мутациях. В результате этого в эволюции скелетные компоненты в целом оказываются менее консервативными, чем мышечные.
При этом целостные морфофункциональные системы (например, скелетно-мышечные комплексы конечностей или челюстного аппарата) проявляют более высокий эволюционный консерватизм, чем отдельные их компоненты (в данном примере кости и мышцы). Это связано с более значительными возможностями авторегуляции систем высокого ранга, в которых авторегуляционные приспособления отдельных элементов дополняются соответствующими свойствами всей системы. Еще Галену было известно, что целый череп прочнее по отношению к ударам, чем его отдельные кости. Причиной этого являются амортизационные свойства шовных соединений между костями.
У многоклеточных организмов авторегуляционные возможности основных морфогенетических систем, обеспечивающих в онтогенезе формирование общего плана строения данной группы, в целом значительно выше, чем таковые генотипа или отдельных фенотипических признаков. Поэтому, как уже было отмечено в главе об эволюции онтогенеза, аппарат наследственности преобразуется в макрофилогенезе значительно быстрее, чем основные фенотипические структуры. Наиболее устойчивы и консервативны в макроэволюции эпигенетические корреляционные системы организма, обладающие наибольшими авторегуляционными возможностями.
Таким образом, корреляционные системы организма, представляющие собой гомеостаты, проявляют тенденцию к эволюционной стабильности. Можно допустить, что нормальное функционирование таких систем происходит в определенных режимах, действующих в пределах основных состояний или типов их конструкции, промежуточные состояния между которыми функционально неэффективны и поэтому эволюционно неустойчивы. Это обусловливает дискретность макроэволюционных преобразований, т.е. их квантовый характер. Макроэволюция представляет собой переход от одного устойчивого состояния морфофункциональных, морфогенетических и генетических систем организмов к другому их устойчивому состоянию.
В целом можно сказать, что характерные пульсирующие темпы макроэволюции, т.е. чередование типогенеза и типостаза в макрофилогенезе, являются следствием основных свойств корреляционных биологических систем, и в первую очередь - целостного организма.
ЗАКЛЮЧЕНИЕ
Подводя итоги, прежде всего вкратце резюмируем основные положения современной эволюционной теории.
Жизнь представляет собой форму существования сложных органических соединений - нуклеопротеидов, структурно организованных в виде биологических систем разного ранга. Структурными единицами живой материи являются интегрированные целостные системы - организмы. Сложность живых систем сама по себе повышает их уязвимость, снижает устойчивость перед лицом изменений внешней среды. Разрушению и гибели живых систем при неблагоприятных изменениях внешних условий противостоят два их важнейших свойства: 1) способность к саморегуляции (системный гомеостаз) и 2) способность к самовоспроизведению, основанная на свойствах молекул нуклеиновых кислот (редупликация) и ведущая к размножению систем данного класса.
Эволюция организмов представляет собой процесс исторических преобразований всех уровней организации биологических систем - от молекулярного до биосферного. Эволюция является неизбежным следствием, вытекающим из основных свойств организмов - размножения и редупликации аппарата наследственности. В изменчивых внешних условиях эти процессы неизбежно сопровождаются возникновением мутаций, поскольку устойчивость любой системы имеет свои пределы. Наличие наследственных вариаций строения и функций организмов при борьбе за существование ведет к возникновению естественного отбора - преимущественного выживания и преимущественного оставления потомства организмами, лучше приспособленными к условиям существования. Результатом действия естественного отбора является приспособительная эволюция организмов. Можно сказать, что эволюция есть форма существования организмов в изменяющейся внешней среде.
Эволюционные изменения разных уровней иерархической организации живых систем (молекулярного - организменного - популяционного) взаимосвязаны, но их связи не имеют простого и однозначного характера. Лабильность этих связей допускает определенную степень независимости и специфику преобразований разных уровней. Естественный отбор непосредственно воздействует на фенотипы целостных организмов в популяциях данного вида. Адаптивные преимущества определенных фенотипов повышают шансы на их выживание и оставление ими потомства по сравнению с хуже приспособленными вариантами. Это приводит к преимущественному воспроизведению соответствующих генотипов и, как следствие, к распространению в популяции генетических комплексов ("супергенов") и отдельных аллелей, обусловливающих развитие адаптивных фенотипических признаков. Результатом этого являются направленные преобразования генофондов популяций, которые представляют собой основное содержание микроэволюционных процессов. При достаточно длительной изоляции разных популяций друг от друга изменения в их генофондах обычно приобретают различные направления, что приводит в конечном итоге к невозможности обмена генетической информацией между ними и тем самым к обособлению изолированных популяций друг от друга в качестве самостоятельных новых видов. Видообразование может происходить как постепенно, в течение длительных сроков, так и сальтационно (при смене немногих поколений) в результате "генетических революций" в популяциях основателей, при полиплоидизации, гибридогенезе, стасигенезе.
Помимо основных факторов эволюции (мутации и отбор) на характер протекания микроэволюционных процессов оказывают влияние различные дополнительные факторы (изоляция популяций, изменения их численности, дрейф генов, молекулярный драйв, гибридизация).
В связи с некоторыми особенностями организации генетического кода (вырожденность, наличие молчащих участков ДНК, повторные последовательности нуклеотидов) и функциональной эквивалентностью ряда аминокислот, на молекулярном уровне эволюции биологических систем происходит более или менее непрерывное накопление относительно нейтральных в адаптивном отношении наследственных изменений, не отражающихся на фе-нотипическом облике и не ведущих к развитию приспособлений организмов ("молекулярная эволюция"). При изоляции друг от друга разных популяций накопление в их генофондах различных изменений может привести к обособлению независимых филетических линий, т. е. к возникновению новых видов, различия между которыми первоначально не имеют приспособительного характера. Однако основным содержанием эволюции организмов является развитие приспособлений к изменениям условий внешней среды, которое может происходить только под воздействием естественного отбора. Сложные приспособления, которые могут функционировать достаточно эффективно лишь в "готовом виде", формируются на основе уже существовавших прежде органов и структур при смене их адаптивной роли в процессе морфофунк-циональной преадаптации. Эволюционное возникновение качественных новообразований основано на дифференциации полимерных и избыточных структур, имеющихся на всех уровнях структурно-функциональной организации живых систем (повторяющиеся последовательности нуклеотидов, дуплицированные гены и целые хромосомы, метамерные структуры фенотипа и т.п.). Все филогенетические преобразования осуществляются на основе микроэволюционных изменений популяционных генофондов. Однако в широкой исторической перспективе макрофилогенеза при формировании организации новых крупных таксонов обнаруживается ряд общих закономерностей, которые не наблюдаются в микроэволюционных процессах и не следуют непосредственно из действия факторов микроэволюции. Таковы необратимость и направленность макроэволюции, поступательный характер морфофизиологического прогресса, чередование периодов типогенеза и типостаза. Макро- и микроэволюция соотносятся друг с другом как высший и низший уровни иерархической системной организации эволюционного процесса: макроэволюционные преобразования складываются из микроэволюционных изменений, но к ним не сводятся - в том же смысле, в котором любая целостная система не сводится просто к сумме своих структурных элементов. Если особенности протекания микроэволюционных процессов определяются свойствами популяций - биологических систем стохастического типа, с нежесткими связями взаимозаменяемых компонентов, то закономерный и направленный характер макроэволюции обусловлен свойствами целостных организмов - биологических систем корреляционного типа с высокой степенью интеграции разнокачественных элементов.
Особенности организации конкретных групп организмов создают предпосылки для развития определенных типов приспособлений и исключают развитие других типов, делают возможными одни направления эволюционных преобразований и запрещают другие. Закономерный поступательный характер эволюции по пути морфофизиологического прогресса определяется отбором на совершенствование механизмов, повышающих независимость организма от внешней среды, при контролируемом самим организмом обмене веществ и энергии со средой. Длительная устойчивость общего плана строения крупных таксонов в периоды типостаза связана с совершенствуемыми стабилизирующим отбором авторегуляционными механизмами в основных системах организмов. Если же преобразования этих систем все же происходят, корреляционные и координационные связи вовлекают в них различные другие системы организма, и вся совокупность этих преобразований развивается за относительно краткие (в масштабах макрофилогенеза) сроки. Высокие темпы типогенеза обусловлены различными организменными факторами, характеризующимися пороговым действием (триггерный эффект изменений ключевых признаков при каскадных взаимодействиях различных систем организма, морфофункциональная преадаптация, дискретные изменения программ онтогенеза и др.).
При этом отбор является основным движущим фактором эволюции, без участия которого невозможна реализация каких бы то ни было потенций развития, обусловленных системными свойствами организмов. Отбор движет эволюцию и придает эволюционным преобразованиям характер приспособлений к изменениям внешней среды, а организменные направляющие факторы определяют конкретные направления и формы происходящих эволюционных перестроек.
Закономерный и направленный характер макроэволюции позволяет поставить вопрос о возможности прогнозирования эволюции. Решение этого вопроса связано с анализом соотношений необходимых и случайных явлений в эволюции организмов. Как известно, в философии категориями необходимости и случайности обозначают разные типы связи явлений. Необходимые связи определяются внутренней структурой взаимодействующих явлений, их сущностью, коренными особенностями. Напротив, случайные связи имеют внешний характер по отношению к данному явлению, будучи обусловлены побочными факторами, не связанными с сущностью этого явления. При этом случайное, конечно, не беспричинно, но его причины лежат вне причинно-следственного ряда, определяющего сущность данного явления. Случайность и необходимость относительны: случайное для одного причинно-следственного ряда является необходимым для другого, и при изменении условий случайные связи могут превратиться в необходимые, и наоборот. Статистическая закономерность представляет собой выявление необходимых, т. е. внутренних, существенных связей среди многочисленных внешних случайных взаимодействий.
Очевидно, надежные прогнозы принципиально возможны для процессов, основанных на необходимых связях компонентов данной системы. С другой стороны, для систем, в которых действуют статистические закономерности (т. е. имеется множество независимых причинно-следственных рядов, но лишь немногие из них имеют существенный, а большинство - случайный характер) возможен лишь вероятностный прогноз. Наконец, прогнозы мало достоверны или практически недостоверны для систем, подчиненных действию нескольких независимых и примерно равнозначных рядов причинно-следственных отношений, поскольку развитие таких систем принимает случайный характер.
Как мы видели выше, возникновение мутаций и естественный отбор определяются основными (сущностными) свойствами живых систем, существующих в изменчивой внешней среде. В свою очередь, эти факторы с необходимостью обусловливают дивергентную и приспособительную эволюцию организмов. Поступательный и закономерный характер эволюции по пути морфофи-зиологического прогресса также является необходимым результатом направляемой отбором эволюции, вытекающим из основных свойств живых систем. Однако он проявляется лишь как общая тенденция в масштабе макроэволюции, т. е. как своего рода статистическая закономерность в филогенезе многочисленных групп организмов, поскольку сказывается влияние многих других причинно-следственных рядов, связанных с изменениями внешней среды. Такой же характер имеют и другие закономерности макроэволюции.
По отношению к развитию жизни на Земле эти выводы сделаны, так сказать, "задним числом" и не имеют прогностического характера. Однако на их основе мы можем с высокой достоверностью предсказать, что после возникновения жизни на любой планете организмы должны претерпеть дивергентную, приспособительную и арогенетическую эволюцию.
Конкретные же направления эволюционных преобразований любой данной группы организмов определяются, с одной стороны, действием отбора (соответствующим характеру изменений внешней среды), с другой - совокупностью организменных направляющих факторов, допускающей возможность некоторого выбора из "разрешенных" вариантов эволюционных изменений ("спектр эволюционных потенций" таксона). Действие отбора имеет статистический характер, а изменения внешней среды лишь отчасти контролируются самими организмами (например, в пределах биоценотической регуляции), но в значительной степени не зависят от них. Таким образом, изменения разных компонентов системы "организм-среда" для каждого вида определяются взаимодействиями многочисленных независимых причинно-следственных рядов, что делает прогнозирование эволюционных преобразований конкретных видов организмов в естественных условиях малодостоверным. Однако при возможности контроля за изменениями условий задача упрощается: при достаточно полном знании организации данного вида теоретически возможно определить спектр его эволюционных потенций и прогнозировать направление эволюционных преобразований при данном направлении изменений условий по крайней мере на вероятностном уровне. Сколько-нибудь полный анализ такого рода еще не осуществлен ни для одного вида организмов и является одной из актуальных задач современной эволюционистики. Наиболее слабым звеном здесь остается недостаточное понимание сущности процессов эмбриональной дифференцировки, лежащих в основе сложных соотношений генотипа и фенотипа и эволюционных преобразований онтогенеза. Это одна из интересных перспектив дальнейшего развития науки.
Что же касается направленного воздействия на виды организмов с целью их изменения в интересах человека, то оно в разных формах на эмпирической основе осуществляется уже давно. Напомним, что практика селекционеров послужила важным пунктом в цепи рассуждений, которые привели Дарвина к открытию естественного отбора. В свою очередь, селекция получила в эволюционной теории научную основу, подкрепленную ныне мощным арсеналом современных методов исследования. Развитие молекулярной биологии в последние годы открыло новые возможности для направленного воздействия непосредственно на наследственную основу организмов с целью получения новых форм: разрабатываемые ныне методы генной инженерии позволяют перестраивать молекулу ДНК с соответствующими изменениями наследственных свойств организмов.
Эволюционистика далеко еще не разрешила всего огромного круга стоящих перед нею проблем и продолжает стремительное развитие. Помимо традиционного обобщения и переосмысливания данных, полученных в области других биологических наук, начинают формироваться ее собственные методы. Среди них следует упомянуть постановку экспериментов на природных популяциях различных видов для изучения действия естественного отбора, внутри- и межвидовых отношений и их эволюционной роли. Аналогичные проблемы решаются и на модельных лабораторных популяциях с использованием методов популяционной генетики. Развиваются методы математического моделирования различных эволюционных процессов. Вероятно, в недалеком будущем важную роль в решении эволюционных проблем будут играть методы генной инженерии и экспериментального вмешательства в онтогенез.
Среди центральных проблем современной теории эволюции следует назвать коэволюцию разных видов в естественных сообществах и эволюцию самих биологических макросистем - биогеоценозов и биосферы в целом. Продолжаются оживленные дискуссии о роли в эволюции нейтральных мутаций и дрейфа генов, о соотношениях адаптивных и неадаптивных эволюционных изменений, о сущности и причинах типогенеза и типостаза в макроэволюции, неравномерности ее темпов, морфофизиологическом прогрессе и т.д. Многое еще предстоит сделать даже в наиболее разработанных областях эволюционистики - таких, как теория отбора, учение о биологическом виде и видообразовании.
Насущной задачей эволюционистики является переосмысливание и интеграция новейших данных и выводов, полученных в последние годы в области молекулярной биологии, онтогенетики и макроэволюции. Некоторые биологи говорят о необходимости "нового синтеза", подчеркивая устарелость классических представлений синтетической теории эволюции, являющейся, в сущности, в основном теорией микроэволюции, и необходимость преодоления характерного для нее узкоредукционистского подхода. Однако в<
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|





